
Chapter 2. What We Know About Large Trees That Fall to the Forest Floor
Chris Maser, Steven P. Cline, Kermit Cromack, Jr.,
James M. Trappe, and Everett Hansen
Summary
This chapter explains how living and dead trees are linked together in the living machinery of a forest.
Introduction
Pacific Northwest forests contain an array of fallen trees in various stages of decay, both in young and old-growth stands, in natural, managed, and recently clearcut stands. Most trees of Douglas-fir region forests are conifers, of which there are six major lowland species north of the Siskiyou Mountains (table 2.1). In total, these temperate forests contain 25 conifer species, many of which are the largest and longest lived of their genera (Waring and Franklin 1979). Large, fallen trees are unique, critical components of forest systems (Franklin and Hemstrom 1981, Franklin and others 1981, Maser and others 1979).
Future forests will contain much less coarse woody debris (CWD), and that debris will be smaller and of different quality than that seen today. We have the technology to remove most coarse woody debris from the forest; in fact, current wood utilization standards encourage such removal (fig. 2.1). Moreover, converting natural forests to intensively manipulated stands reduces tree lifespans from centuries to decades; future trees will be much smaller than they are today, and wood quality will undoubtedly be different from that of today’s forests.
Individual trees have different characteristics that produce diversity within the forest. Trees can live for hundreds or even thousands of years because they produce new functional tissues every year, but most die much earlier from injury, disease, or both.
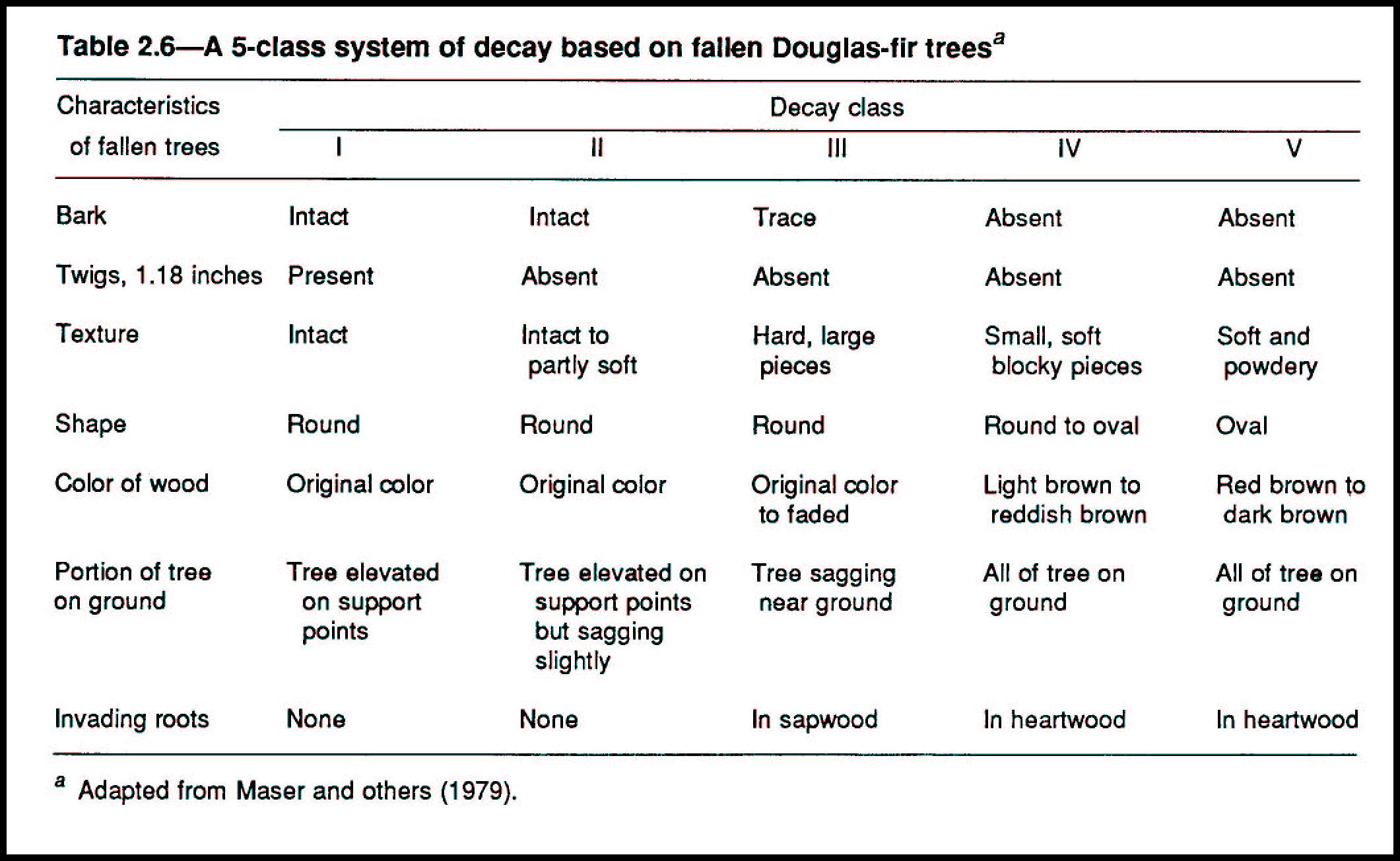
The cause of death (table 2.2) determines diversity of the structural and functional roles served by the dead tree in its tenure on the forest floor. When a tree dies and remains standing as a snag, this new, different structural and functional unit adds diversity to the system, diversity important to many forest organisms (fig. 2.2; tables 2.3, 2.4, 2.5). When a snag or a green tree falls to the forest floor, diversity again is added (figs. 2.1, 2.3, 2.4; tables 2.6, 2.7).
Page 25

Figure 2.1-A clearcut on Marys Peak in which most of the large woody debris has been removed. Note how small the remaining pieces are.
Page 26

Sources: Aho (1966, 1982); Bull and others (1980); Crouch (1969); Etheridge (1973); Hadfield (1985); Hawksworth and Wiens (1972); Houston (1981); Johnson and others (1964); Koonce and Roth (1980); Krebill (1968); Leaphart (1959); Maser (1967); Neff (1928); Poelker and Hartwell (1973); Roth (1970); Ruth and Yoder (1953); Sanders (1964); Schowalter (1985); Shigo (1973, 1979) Shigo and Marx (1977); Shortle (1979); Wargo (1984); Wright and Harvey (1967).

Figure 2.2-Snags are important structural components of the forested landscape.
Page 27

Page 28

Page 29

Page 30
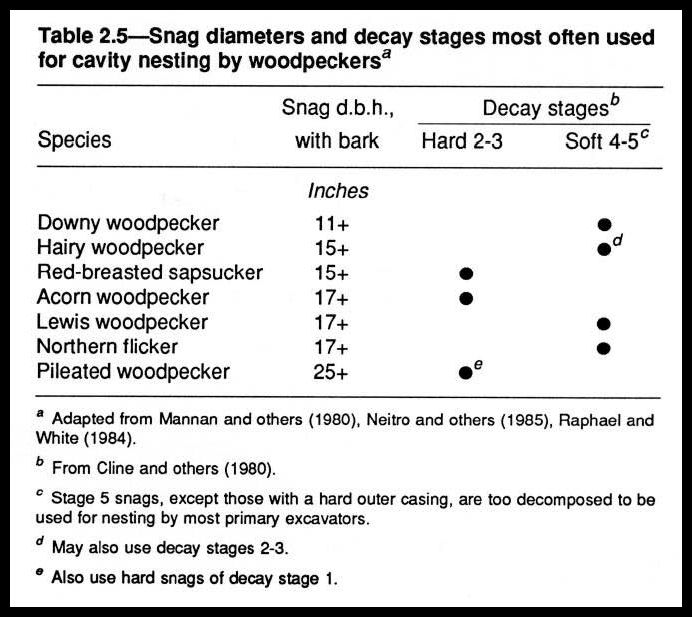

Figure 2.3-Fallen trees add a critical dimension of diversity to the forest floor. The man is 6 feet 7 Inches tall.
Page 31

Page 32
Forest Floor Diversity
The forest floor is the interface between the belowground and the aboveground components of the forest. As a center of intense physical, chemical, and biotic activity, it both influences and reflects ecosystem functions. In turn, ecosystem functions reflect the array and quality of processes resulting from structural diversity.
Physical Diversity
When a tree falls-be it a wind-uprooted, healthy tree or a decomposing snag-a sequence of ecological consequences begins. The fall of understory trees, usually western hemlock or western redcedar, is caused by wind being funneled below the overstory canopy. Falling of understory trees does not visibly affect the overstory canopy, but additional light is admitted to the forest floor. The falling of an overstory Douglas-fir, however, creates an evident canopy opening in which shade-tolerant understory trees will be released to grow.
The opening increases considerably if the falling tree causes successive uprooting and breaking of neighboring trees. The probability of a tree causing such a domino effect when it falls increases with its size and vigor. A large, healthy tree is more likely to fell the trees it strikes than is a decayed snag, which is more likely to break or shatter. A large snag falling in a young stand is more likely to knock over a smaller live tree than is one falling in a mature or old-growth stand.
The effect on canopy space is less dramatic when a tree dies standing and becomes a snag. The snag occupies space and casts shade that affects the understory vegetation. Over time, however, the snag deteriorates, its effect changing as decomposition advances and pieces slough off.

Figure 2.5-Wind may break unhealthy trees above the ground.
Fallen trees create opportunities for new plants to become established. An old-growth Douglas-fir may be uprooted or may break off at or above the ground, depending on its health (fig. 2.5, color). Trees that break near the ground are infected with butt or stem rots. The roots and stump decompose in place, leaving understory vegetation intact and creating few new sites for establishment of vegetation. A tree with root rot may be uprooted, but the roots break off instead of being pulled from the soil.
Page 33

Figure 2.6-Wind blows over healthy trees and pulls their roots from the soil.
In contrast, when roots of a healthy tree are pulled out of the ground when it falls, bare soil is exposed as an extensive root-pit and mound (fig. 2.6, color). The bare mineral soil of a root-pit and mound-and with time the fallen tree itself presents habitats readily colonized by tree seedlings and other plants (Christy and others 1982).

Figure 2.7-Forest floor diversity is partly maintained by windthrown trees that create a pit-and-mound topography as they are uprooted.
The surface of a forest floor is roughened by fallen tree stems, their uprooted butts, and the pits and mounds left after the uprooting (fig. 2.7, color). Living trees roughen the surface of the forest floor by sending roots outward along slopes, often near the litter layer. Tree stems also affect the surface by sloughing bark, which arrests soil creep at their bases. Of all the factors affecting surface microtopography, the pit-and-mound topography produced by uprooting trees is best known (Beatty 1984, Bratton 1976). Topography is a major influence in creating and maintaining species richness of the herbaceous understory (Bratton 1976, Thompson 1980) and on the success of tree regeneration (Henry and Swan 1974). Pit-and-mound topography is common in all types of forests and is a major cause of soil mixing (Stephens 1956).
Live tree roots increase soil shear strength; as dead roots decompose, soil strength declines. For this reason, mass soil movement on slopes increases after clearcutting (Burroughs and Thomas 1977, O'Loughlin 1974, Ziemer 1981).

Figure 2.6-Large stumps from old-growth trees are a finite resource, and their loss from the forest affects both soil shear strength and watershed hydrology.
As tree stumps and roots decompose, they often form interconnected, surface-to-bedrock channels that rapidly drain excess water (Beasley 1976, de Vries and Chow 1978, Gaiser 1952) (fig. 2.8, color). The collapse and plugging of these channels during latter stages of decomposition are believed to foster slope instability after logging by forcing more runoff to drain through the soil matrix, reducing soil cohesion and increasing hydraulic pressure (Chamberlin 1972).

Figure 2.9-A fallen tree oriented along the contour of a slope. The upslope side is filled with humus and inorganic material that allows invertebrates and small vertebrates to tunnel alongside. The downslope side provides protective cover for larger vertebrates. When under a closed canopy, such trees are also saturated with water and act as a reservoir during the dry part of the year.
Large, stable tree stems lying along slope contours reduce erosion by forming a barrier to downhill soil movement (fig. 2.9, color). Soil and forest floor materials deposited along the upslope side of a fallen tree reduce nutrient loss and form an excellent site for establishment and growth of vegetation.
Chemical Diversity
Fallen trees are a major source of soil organic material in Pacific Northwest forests. They also contain essential nutrients that are released during decomposition by micro-organisms and plants, fungi, and animals, such as termites (fig. 2.10), that derive nutrients from the micro-organisms. The decomposer community is adapted to the relatively low nutrient concentrations in wood; wood-decomposing fungi can survive and function at nitrogen concentrations about one-third those usually found in undecomposed wood (Cowling and Merrill 1966).
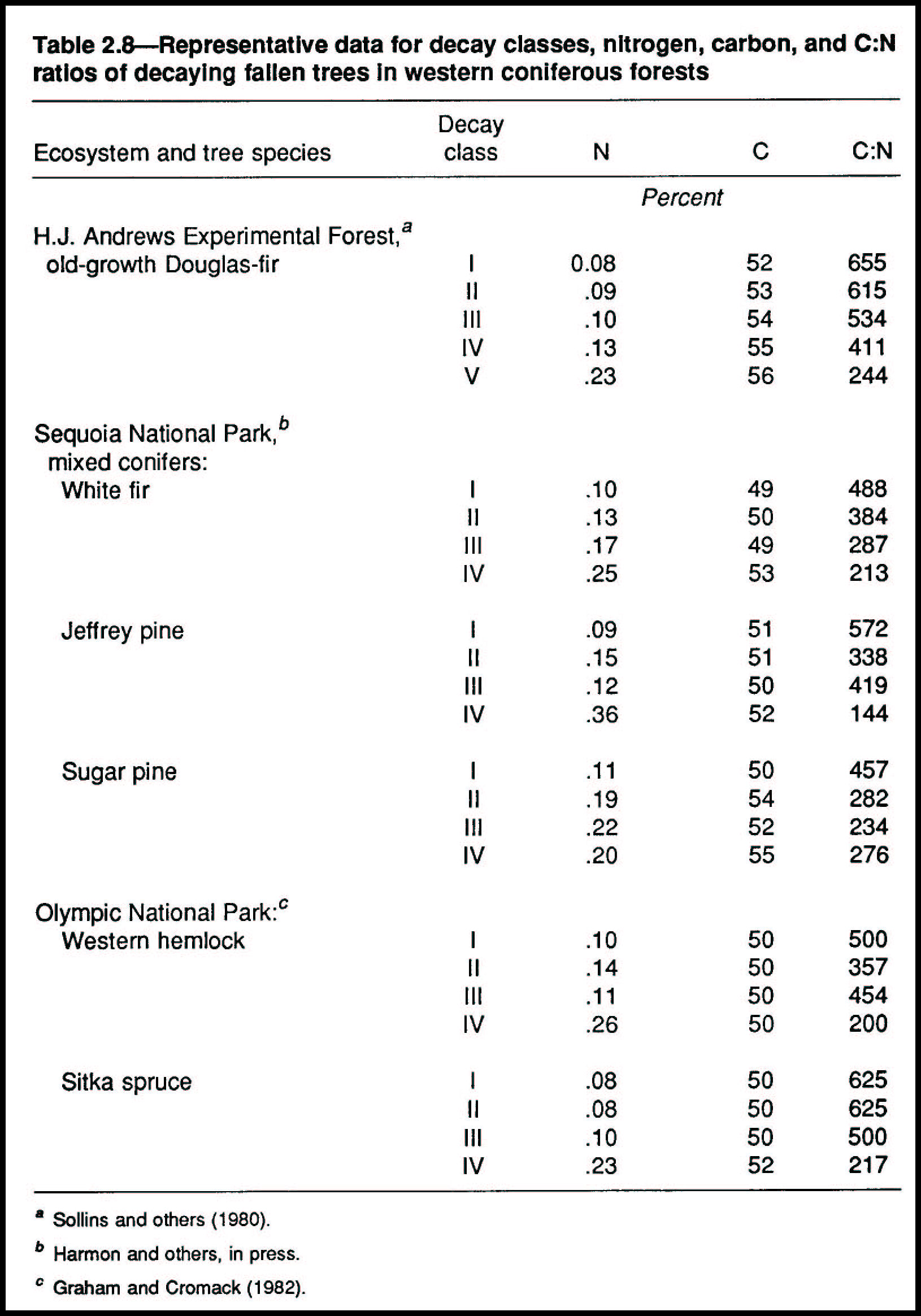
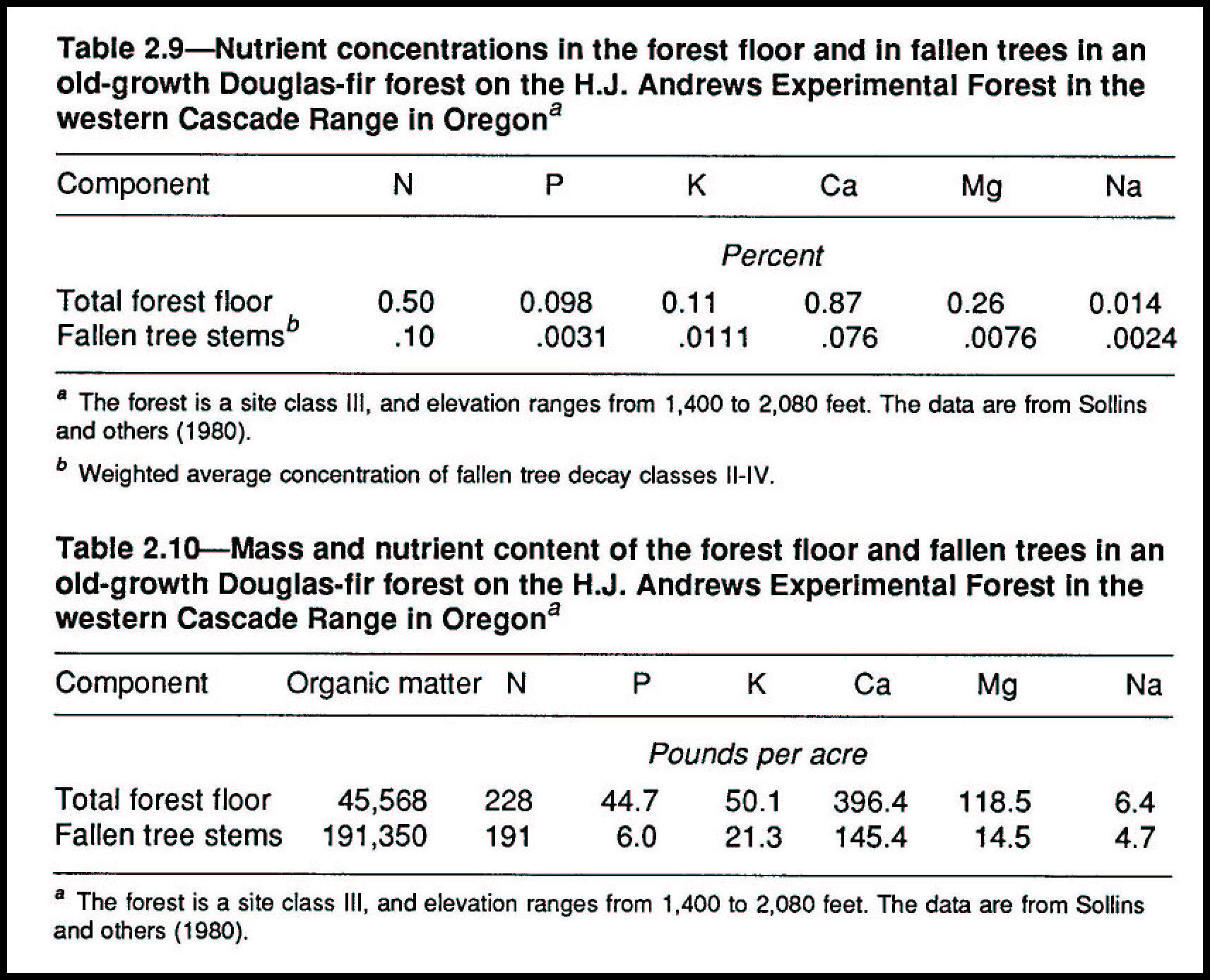
The five decay classes (table 2.6) of fallen trees have different concentrations of nutrients-the higher the decay class, the higher the nutrient concentration (tables 2.8 and 2.9). As carbon compounds in wood decay to carbon dioxide, water, and residual compounds, their nutrient contents, such as nitrogen, are released from the wood matrix. Micro-organisms and the larger creatures that decompose the wood for energy and structure concentrate these nutrients in their tissues as they metabolize the carbon compounds.
Page 34

Figure 2.10-A Pacific dampwood termite worker. A termite can digest the wood it eats because of a mutually beneficial, three-way relation between the individual termite, cellulose-digesting protozoa,, and nitrogen-fixing bacteria that live in its gut. The protozoa digest the cellulose in the wood and convert it to a form that the termite can use as food. In turn, the termite’s body provides an anaerobic chamber that is replete with food for the protozoa. Wood particles eaten by the termite pass into the chamber and are engulfed by the protozoa, which in turn ferment the cellulose. Major products of fermentation are carbon dioxide,, hydrogen, and acetic acid. The acetic acid is absorbed through the wall of the termite’s hindgut and is oxidized as energy (Breznak 1975).
Nitrogen concentrations can increase several fold during progressive stages of decay (table 2.8), in part because of biological nitrogen fixation by free-living bacteria in the wood (Silvester and others 1982). Absolute gains or losses of nitrogen or other elements depend on the balance between nutrient inputs and losses during decay. Nutrients that are conserved by plants and animals during decomposition include nitrogen, calcium, magnesium, and sodium. Others, such as potassium, usually are leached and recycled (Grier 1978, Yavitt and Fahey 1982).
Fallen trees are a substantial reservoir of soil organic matter in old-growth Douglas-fir forests (table 2.10). In two Douglas-fir forests, the organic matter from all decay classes of fallen trees older than 500 years represented about 30 percent of the total soil organic matter (Means, Cromack, and MacMillan, unpublished data). Nutrients are less concentrated in fallen trees in all decay classes than in leaf, twig, and root components of the forest floor (table 2.9). Because of the great amount of large wood in an old-growth Douglas-fir forest, however, the total amount of nutrients in fallen tree stems is still important (table 2.10). The proportion of the total soil organic matter contributed by fallen trees can be nearly fourfold greater than that of the other forest floor materials (table 2.10). Litter components take 10-50 years to decompose (Fogel and Cromack 1977), whereas fallen trees take as long as several centuries (Means and others 1985). Thus, the nutrient pool of a fallen tree cycles on a much longer time scale than that of the rest of the forest floor.
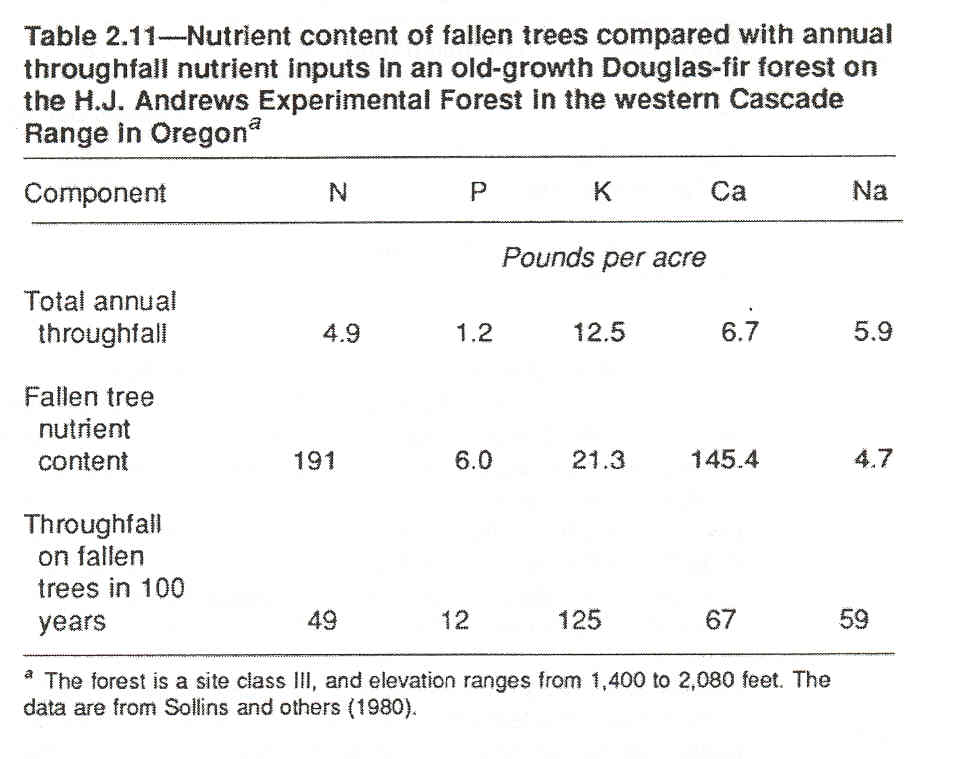
The forest floor and its decaying wood receive nutrients from throughfall (rain, snow, or dew that picks up nutrients as it falls through the canopy) (fig. 2.11). If fallen trees occupy, say, 10 percent of the ground surface in an old-growth Douglas-fir forest, and if they remain in place for 100 years, they potentially could accumulate nutrients equivalent to 10 times the nutrients carried in the average yearly throughfall (table 2.11).
Page 35
Page 38
Decomposition of fallen trees is aided by a variety of micro- and meso-organisms that live in, on, or near them (Elton 1966, Harmon and others 1986). Potential thus exists for considerable uptake, utilization, and export of nutrients from fallen trees during decomposition. Substantial cycling of nutrients occurs, for example, in decayed, fallen lodgepole pine (Yavitt and Fahey 1985), especially after snowmelt. Decomposition of fallen trees releases nutrients for microbial and plant growth (Yavitt and Fahey 1985). Fruiting bodies of hypogeous mycorrhizal fungi produced in association with tree roots in rotten wood provide nutrients and energy to insects, mollusks, and mammals (Maser and others 1979).
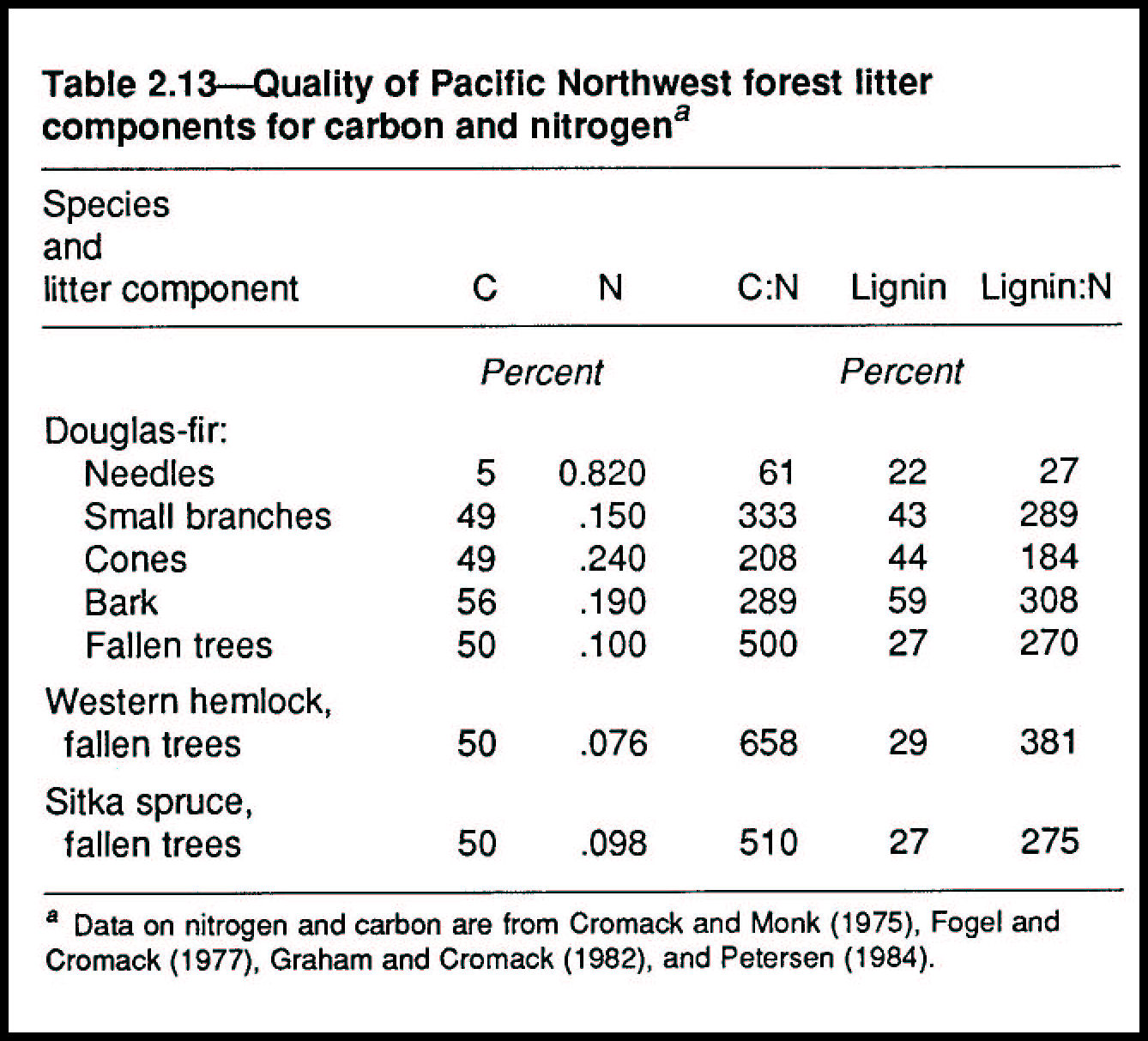
Woody litter, regardless of type or size, takes considerably longer to decompose than does needle and leaf litter (table 2.12). Needles, leaves, and small twigs decompose faster than larger woody material and nutrients are thereby recycled faster in the forest floor. About 140 years are needed for nutrients to cycle in large, fallen trees and more than 400 years for such trees to become incorporated into the forest floor; they therefore interact with the plants and animals of the forest floor and soil over a long period of forest and stand successional history (table 2.12). During this time, a variety of shrubs and trees, such as western hemlock and Sitka spruce, have the opportunity to develop part or all of their root systems within rotting wood (Harmon and others 1986).
Page 37

Figure 2.11-Large fallen trees decay slowly and over time receive nutrients from such ecological processes as rain falling through a nutrient-rich canopy.
Release of nutrients, especially nitrogen, from forest litter is a function of the decomposition rates and the ability of micro-organisms to retain the nitrogen in their tissues (Mahendrappa and others 1986). Work of Bosatta and Staff (1982) suggests that, for slower decomposing substrates, such as wood, the rate of nitrogen release per unit of carbon mineralized increases as the decomposition rate decreases. Nitrogen concentration is an important variable in the regulation of decomposition, especially in the earlier stages of decay. Lignin is important in later decomposition because it affects the proportions of different residues that may be incorporated into humic materials (Berg 1986) (table 2.13).
Page 38

Page 39
Woody litter components are generally higher in initial lignin than are nonwoody components (table 2.13); high lignin content results in formation of large quantities of humus in the latter stages of decay (Berg 1986; Berg and McClaugherty, in press; de Hann 1977). High concentrations of initial lignin and higher lignin-to-nitrogen ratios in woody substrates mean they decompose slowly (Melillo and others 1982).
Humus formation is important in regulating the incorporation of nitrogen into humic materials (Allison 1973, Broadbent 1970, Mahendrappa and others 1986, Russell 1973). Because of its high cation exchange capacity and slow decomposition, rotten wood can retain available mineral nitrogen from throughfall and decomposition as well as organic nitrogen compounds mineralized within the wood chemical matrix. Roots and mycorrhizae of plant species that colonize decaying wood use its available nitrogen.
Animals facilitate the process of microbial colonization of fallen trees by such activities as boring and tunneling (Elton 1966, Maser and Trappe 1984a). Ants and termites excavate 10-18 percent of the fallen trees, such as Jeffrey pine and sugar pine, in Sequoia National Park, California (Harmon and others, in press). Ants also extensively excavate decaying mountain hemlock near Waldo Lake, west-central Oregon Cascades (Cromack, unpubl. data). In addition to excavating, termites have internal nitrogen-fixing micro-organisms in their intestines (Breznak 1975, Elton 1966).
Nitrogen limits forest productivity in the Pacific Northwest, so it is important that decaying, fallen trees are sites of nitrogen fixation by free-living bacteria (Cornaby and Waide 1973, Silvester and others 1982). The rates of nitrogen fixation in decaying wood are relatively low compared with those of symbiotic nitrogen-fixing plants, such as red alder and snowbush (Binkley and others 1984, McNabb and Cromack 1983, Tarrant and Miller 1963, Youngberg and Wollum 1976). About 1 pound per acre per year of nitrogen is fixed in decaying wood (Sollins and others 1986). About half the volume of the annual input of nitrogen is in rainfall (Sollins and others 1980).
The long-term input by nitrogen fixation in decaying fallen trees and by canopy inhabiting lichens maintains a positive balance of nitrogen in the ecosystem. Little nitrogen is lost through soil erosion and leaching in undisturbed forests, so additions through relatively small rates of nitrogen fixation and input from precipitation gradually accumulate to significant amounts. This positive balance permits periodic, large losses of nitrogen by fire and accelerated erosion to be more than recovered when added to large inputs of belowground nitrogen fixation from early successional, nodulated species, such as red alder and snowbush.
The net gains of nitrogen through time have led to substantial accretion of nitrogen and associated soil organic matter in many undisturbed forests of the Pacific Northwest. Nitrogen in these forests has accumulated from about 17,800 pounds/acre on highly productive sites to 2,670-4,450 pounds/acre on poor sites. Even forests low in nitrogen, such as a mountain hemlock forest growing on tephra deposited by the eruption of Mount Mazama about 6,700 years ago, has gained 1,335-1 ,602 pounds/acre of nitrogen in the absence of major nitrogen-fixing plants (Boone 1983). This gain may be due in part to the presence of coarse woody debris and some nitrogen-fixing lichens.
Page 40
Classical agricultural soil theory suggests that nitrogen is available to plants only when the C:N ratio is less than about 25:1 (Russell 1973). Even in advanced decay, the C:N ratio in fallen trees remains consistently much higher than that of the forest floor. An important new hypothesis, however, suggests that nitrogen availability is controlled as much by the availability of easily decayed carbon substrates as by nitrogen concentration per se (McClaugherty and Berg, in press). Wood-rotting fungi can extract nitrogen from substrates with a C:N ratio of 1800:1 (table 2.8), nearly three times as much as sound wood of typical western conifer species. Mycorrhizal fungi, the major organs of nutrient uptake by the roots, also have decomposing capability (Durall, Todd, and Trappe, unpubl. data). Thus, decaying wood has long-term potential for contributing nitrogen for tree growth as residual lignin and humus are decomposed.
The new theory regarding nitrogen availability suggests that decomposing wood loses easily biodegradable carbon substrates faster than do other litter components, such as leaves (Berg 1986; McClaugherty and Berg, in press). Consequently, nitrogen from decaying wood becomes available for plant growth at the higher C:N ratios.
Wood appears to facilitate colonization by long-lived decomposer and mycorrhizal fungi that capture nutrients, such as nitrogen, within a chemical matrix that promotes nitrogen retention. A slow-release nitrogen system operates in coarse woody debris because nitrogen has the potential for becoming available for plant growth at higher C:N ratios than in typical agricultural residues (Berg 1986; McClaugherty and Berg, in press). Other beneficial properties of large, fallen trees in forests include high waterholding capacity and cation-exchange capacity (Means and others 1985, Sollins and others 1986), and habitat for micro-organisms, invertebrates, vertebrates, and higher plants (Maser and Trappe 1984a) (fig. 2.9).
Biotic Diversity
The manner in which a fallen tree comes to rest on the forest floor greatly influences subsequent diversity of both external and internal plant and animal habitats. The decomposing fallen tree provides a changing spectrum of habitats over many decades-even centuries. It provides diversity within a given successional stage and forms a physical-chemical link through the many successional stages of a forest.
Placement of fallen trees-how they lie on the landscape determines the chain of events leading to diversity. The cause of tree death and characteristics of the site and its forest stand mainly determine the initial placement of a fallen tree (fig. 2.12). In the absence of human interference, the ultimate placement is determined by how it decomposes and becomes incorporated into the soil (Franklin and others 1981,Maser and Trappe 1984a).
A fallen tree performs various ecological functions between the time it falls and the time it is finally incorporated into the soil. If it lies up-and-down slope or falls across other downed trees, most of its volume is initially suspended above the ground. Such elevated relief adds complexity to the forest floor by creating cover and shade.
Page 41

Figure 2.12-Diversity on the forest floor is created by the way trees fall; here a Douglas-fir tree, decay class I, fell before the Douglas-fir snag, which entered decay class III.
When a fallen tree decomposes, unique new habitats are created within its body as the outer and inner bark, sapwood, and heartwood decompose at different rates. The quality of the habitat depends on its physical and chemical properties (fig. 2.13).
A fallen tree interacts with its environment through internal surface areas. A newly fallen tree is not yet a habitat for plants or most animals. But once organisms gain entrance to the interior, they consume and break down wood cells and fibers. Larger organisms-mites, collembolans, spiders, millipeds, centipedes, amphibians, and small mammals-must await the creation of internal spaces before they can enter. The flow of plant and animal populations, air, water, and nutrients between a fallen tree and its surroundings increases as decomposition continues (fig. 2.14).
Internal space develops in a fallen tree through physical and biological processes. A tree cracks and splits when it falls and dries. Microbial decomposition breaks down the cell wall and further weakens the wood. Wood-boring beetle larvae and termites tunnel through the bark and wood, not only inoculating the substrates with microbes but also opening the tree to colonization by other microbes and small invertebrates. Wood-rotting fungi produce zones of weakness, especially between annual rings, by decaying spring wood faster than summer wood. Plant roots that penetrate the decayed wood split and compress it as the roots elongate and grow in diameter. Because of all this internal activity, the longer a fallen tree rests on the forest floor, the greater the development of the surface area of its internal cavities. Most internal surface area results from biological activity, the cumulative effects of which not only increase through time but also act synergistically-insect activity promotes decomposition through microbial activity that encourages establishment and rooting of plants.
Page 42


Page 43
Conclusions
Numerous physical and chemical changes occur as fallen trees decay: (1) density decreases; (2) water content increases until decay classes III and IV are reached, at which time the water content stabilizes (fig. 2.15); (3) mineral and nitrogen contents increase; (4) cellulose content decreases; (5) relative lignin content increases; (6) C:N ratio decreases (Fogel and Cromack 1977, Foster and Lang 1982, Graham and Cromack 1982, Grier 1978, Lambert and others 1980, Sollins 1982); and (7) internal temperature fluctuations are buffered as the fallen tree comes in contact with the ground (Sollins and Cline, unpubl. data).
Habitats, both external and internal, are influenced by tree size. An uninterrupted supply of new, immature wood in young forests decomposes and recycles nutrients and energy rapidly. Habitats provided by the death of young trees are short lived and rapidly changing. In contrast, the less frequent, more irregular mortality of large trees in old forests is analogous to slow-release fertilization. The lasting quality of large fallen trees creates stable habitats in which large woody debris accumulates (fig. 2.76, color). Scattered accumulations of large woody debris are associated with openings in the forest canopy (Stewart 1984). Large fallen trees in such an area often contact each other physically, creating external habitats of intense biological activity (Maser and others 1979, Neitro and others 1985, Stewart 1984, Thornburgh 1967).

Figure 2.16-Large fallen trees are a finite resource that create a myriad of changing habitats through time as they decompose and recycle into the forest soil and new, living trees.
Conclusions
Decaying, fallen trees contribute to long-term accumulation of soil organic matter, partly because the carbon constituents of well-decayed wood are 80-90 percent residual lignin and humus (Means and others 1985). Decaying wood in the soil and establishment of conifer seedlings and mycorrhizal fungi on dry sites are positively correlated (Harvey and others 1987). Fallen trees also create and maintain diversity in forest communities. Soil properties of pits and mounds differ from those of surrounding soil (Beatty and Stone 1985); such chemical and topographic diversity in turn affects forest regeneration processes (Lyford and MacLean 1966). All this, especially large fallen trees that reside on the forest floor for long periods, adds to spatial, chemical, and biotic diversity of forest soils, and to the processes that maintain long-term forest productivity.

Figure 2.15-Water content of fallen Douglas-fir trees in winter and summer; each point represents a decay class from I (newly fallen trees) to V (fallen trees decomposed to a mound of brown cubical rotted wood).
Page 44
Had color figs which were placed in proper place. So this page is blank.
Page 45
This page was intentionally left blank.
Page 46