
Chapter 4. What We Know About Large Trees in Estuaries, in the Sea, and on Coastal Beaches
Jefferson J. Gonor, James R. Sedell, and Patricia A. Benner
Summary
Coarse woody debris is an important part of estuarine and oceanic habitats, from upper tidewater of coastal rivers to the open ocean surface and the deep sea floor.
Introduction
An estuary is a body of water at the coastal end of a river system, in which freshwater mixes with saltwater before it enters the ocean. An appreciably wider shallow basin or embayment with extensive tidelands is in the estuarine section of most Pacific Northwest river systems. In some systems with large flows and narrow, deep channels, such as the Rogue River in Oregon, the truly estuarine region is confined to a short reach upstream of the river mouth. Fallen trees in streams or within reach of flood or tidal waters move downstream to enter estuarine and marine systems.
The estuarine portion of the ecosystem supports important environments and habitats including those of people, most of whom live around the lower ends of river systems. The ecologic and economic role of woody debris in these ecologically complex, productive systems is not well known.
Origin and History of Pacific Northwest Estuaries
During the late Pleistocene, eustatic sea level was about 325 feet lower than it is today. Pacific Northwest coasts were not glaciated, and rivers excavated deep valleys in the flat, exposed coastal plain. Sea levels rose after the close of the Pleistocene, and the coastal plain was submerged to form the present Continental Shelf. The seaward ends of many of these river valleys became flooded, and sediments accumulated in former meanders in the rivers’ old flood plains. The main channels of these drowned rivers remain as the central, deeper channels of modern estuaries that retain the elongate, sinuous topography of a river in its flood plain valley. Channels are relatively shallow for most of their length.
The position and configuration of estuaries, no older than 10,000 to 15,000 years, remain dynamic, but the physiography has changed. Estuaries have long existed on the continental border of the Pacific Northwest, and they have moved across the Continental Shelf and present coastal plain in response to fluctuations in sea level changes.
Today’s estuary continues to accumulate sediments in the former channels and on the flood plains, usually river-derived silts and muds in the upper reaches and increasingly sandy material near the mouth. Estuarine tidal flats and marshes occupy the former flood plain areas along the drowned river valleys that are now estuarine embayments (fig. 4.1).
Page 83
Figure 4.1--An Oregon estuary: Coquille River marsh at Bandon, 1985 (photo, courtesy of Tony Howell).

Figure 4.2-Lower estuary, Tillamook, Oregon, 1871 (photo, courtesy of the Oregon Historical Society, negative ORH 52608).
History of Wood in Northwest Estuaries and Coastal Beaches
U.S. Government reports and early visitors’ journals of the Pacific Northwest documented great amounts of large wood in the estuaries and on the beaches at river mouths. These mid-1800 accounts describe the quantities and size of the drift trees, also called "snags," that significantly exceeded present amounts of woody debris in the lower portions of river systems and beaches (fig. 4.2).
Coast survey reports in the 1850’s recorded that many of the drift trees in the lower Columbia River were as large as 150 feet long by 13 to 18 feet in circumference; the largest was 267 feet long (Secretary of the Treasury 1859). Swan (1971) also reported drift trees as large as 250 feet long by 8 feet at the base, with a root span of some 20 feet, on the beach near the mouth of the Quillayute River in the Washington territory.
Page 84
The U.S. Army Corps of Engineers reported that Northwest estuarine shorelines and river-mouth beaches had often been covered with driftwood in the 1870’s, as was the mouth of the Coquille River in the 1880’s (fig. 4.3).
For several years after the coastal areas were settled in the 1850’s and 1860’s, roads were limited and land travel was impractical, especially in the winter. The coastal rivers were under the influence of tides for 12 to 40 miles from their mouths and had a low-gradient, deep channel along which commercial boats and log rafts could travel. Slowing currents and stormwind patterns, however, created zones of wood deposition in the estuaries. Many snags and sunken driftwood presented major
obstacles for river traffic. The Corps’ responsibility on many rivers during the late 1800’s was to improve and maintain the navigability of the portions of the rivers deemed to be economically important (table 4.1).
Page 85
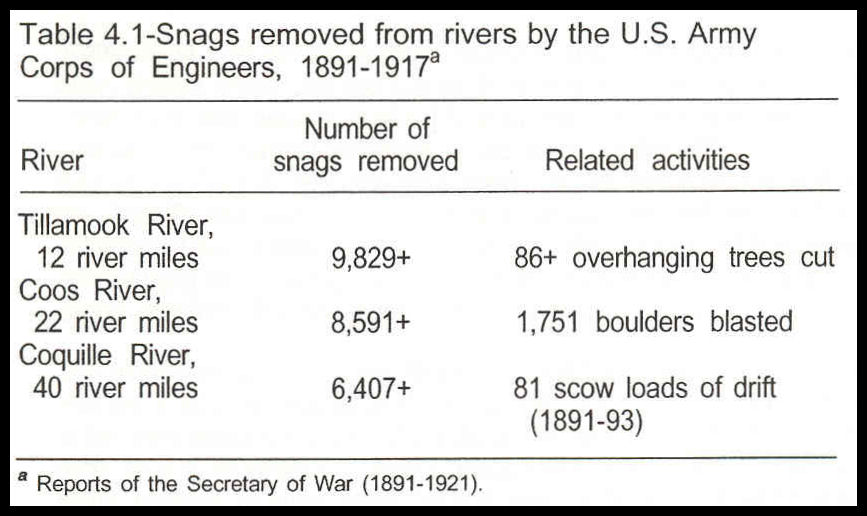
The wood-removal operations by the Corps represented only a portion of the total wood pulled from the lower river systems. Gill-net fishers formed teams to remove wood that threatened to tear fishing nets. Local landowners and later, port authorities, also worked to maintain channel navigability.
Sources of Wood For Estuaries and Beaches
Driftwood is found throughout an estuary, both in the channel and on mudflats and higher vegetated areas. Currently, the greatest concentration of woody debris is found along the shoreline of an estuary out of reach of tidal water. The most stationary wood is in the marshes, or at least partly buried in the channel.
Driftwood deposited in marshy areas between the main channel and the shoreline in the lower Nehalem estuary is estimated to have 50 percent fewer pieces and 60 percent less volume than in 1939. Stranded wood in the marsh in 1939 probably included many escaped lumber company logs that had been floated downriver or held in booms.
Another human-related source of wood is the many pilings and associated wood structures. Port authorities estimated that nine estuaries in Oregon in 1986 had at least 15,000 maintained pilings and thousands more wood structures-old stubs from abandoned log pens and channel projects, old mill pilings, and remnant docks. This wood can be considered a partial substitute for the snags that naturally occurred before the late 1800’s.
Streams and rivers have long supplied estuaries and beaches with wood, and most of the material was transported downstream during higher flows. A captain in the U.S. Army Corps of Engineers in 1897 noted of the Coquille estuary, "It has always been difficult to keep this portion of the river open. The drainage area is densely wooded, and every freshet brings down many stumps, logs and trees; these lodge and arrest sediment, forming shoals." (Report of the Secretary of War 1894-95).
Page 86
Watersheds annually replenished wood in the lower portions of the river basin and often floated wood into the ocean, from which it washed up onto the beaches. The lower river and estuary banks (riparian corridor) probably were the most common sources of the largest driftwood in the bays. In the 1960's, the banks of the upper half of the Coquille estuary were lined with mature hardwoods that made travel on the Coquille like walking "dim aisles in ancient cathedrals" (Dodge 1898). In the Tillamook River system in 1904, the U.S. Army Corps of Engineers cut down all overhanging trees along the banks of the estuary in an attempt to alleviate the woody debris problem (Report of the Secretary of War 1904-5).
The woody vegetation along many river corridors was cut in the 1800’s to clear land and for a local source of wood. Upstream, the riverside forests were among the first to be commercially harvested because the logs could be floated down the river to the ports at a time when no other transportation was available (Sedell and Duval 1985). Major sources of large wood for estuaries and beaches along the Northwest coast were exhausted by 1920.
The ocean is another source of driftwood in estuaries and on beaches. Winter storms blow ocean-transported wood into river mouths and onto coastal beaches, generally north of the debris’ origin. Some of the woody debris may be buried for long periods by river-bottom sediments in the estuaries or in sandy spits on the coast, but much of the wood probably remains fairly mobile. Other driftwood is deposited on the marshes and along the higher ground of the estuary boundaries, where it remains until it decomposes.
Flow Regimes in Pacific Northwest Coastal Estuaries
Estuarine tidal exchange and streamflow features govern the transport and retention time of woody debris and create salinity gradients. This physical regime in turn affects the distribution of marine wood-boring animals and their degradation of wood in these estuaries.
Tide affects river level much farther upstream than the penetration of even the most dilute seawater at the highest tides. This hydraulic effect causes river levels to rise and fall and can even reverse the direction of flow in the reach referred to as the "tidal river." Its farthest upstream extent is the "head of tide." This effect, present throughout the year, increases the rate of transport of floating woody debris faster than might be expected and increases the chance that wood stranded on the shoreline by freshets will be reached by subsequently higher tides and be returned to the downstream flow. Tidal reaches for Oregon rivers vary; the two longest tidally influenced river sections are those of the Columbia (140 miles) and the Coquille (40 miles) (table 4.2).
The most distinctive environmental features of estuaries along their length are the horizontally created gradients of salinity and temperature caused by the mixing of fresh river water with ocean saltwater; such gradients are vertical at times, going from fresh to saline conditions as depth increases. The salinity gradient along the length of an estuary varies greatly over time because of the twice-daily ebb and flow of the tides and the seasonal changes, such as winter floods’ moving the freshwater seaward. These variations are greatest in the middle of the estuary where salinity conditions are intermediate between the upper freshwater and lower saltwater.
Page 87

Seasonal Stratification and Flushing
The density differences between freshwater from streamflow entering an estuary and seawater entering from the mouth can result in vertical stratification of the estuatine water mass. In a density-stratified estuary, a low salinity, low density, mixed surface layer flows toward the sea, and the denser, more saline, seawater originating at the mouth flows upstream near the bottom. This bottom flow replaces the seawater that is entrained and mixed upward, along the length of the estuary, into the surface outflow. This upward mixing continually increases both the volume and salinity of the upper layer as it flows down the estuary.
The presence and extent of vertical-density stratification vary seasonally in Pacific Northwest estuaries, mainly during the rainy winter season when streamflow dominates estuarine hydrography. Winter floods may produce vertical stratification even in the lower segment of the estuary near the mouth; at this season, the upstream flow of saline, ocean water is strong near the bottom and the downstream flow of dilute, mostly riverine water is strong at the surface. The near-bottom flow of marine water weakens gradually in the upstream direction and finally ceases at some point upstream. Flows in the upper estuary above this point are downstream near both the bottom and the surface.
The "null point" in the system in short-term subsurface transport of water and suspended materials, including woody debris, is in the midestuarine region where near-bottom flow is not appreciable either upstream or downstream. Its exact position varies with the volume of river discharge. Below this null point, water and sediment may be transported upstream along the estuary bottom. At and above it is a region of increased turbidity and sedimentation as suspended riverine sediments enter a region of decreased velocity and flocculation. Woody debris may be incorporated into sediments in the estuarine channels of this zone, to be periodically remobilized and flushed farther down the estuary during periodic winter floods when increased
Page 88
rivet-flow moves the null point downstream. In summer, floating wood might be retained in the upper estuary, moving downstream only during tidal cycles with the greatest vertical range. In winter, the hydrographic regime favors downstream transport along the entire estuary.
Water in Pacific Northwest estuaries is not strongly density stratified in summer when seasonal rainfall and resulting streamflow are at a minimum and tidal exchange dominates the estuarine hydrographic regime. The relatively great tidal range of the region results in enough volumes and energy to mix the estuary vertically through most of its length. In summer, difference between surface and bottom water at any point is much less than in winter.
Pacific Northwest estuaries typically have a large proportion of tidelands-to-subtidal area that results in a high exchange ratio (Johnson and Gonor 1982). This is the ratio of the tidal prism (intertidal volume) to the total high tide volume of the estuary and is proportional to flushing time if flow is not restricted. Many of the known flushing times are fewer than 10 days in summer. Because of the length of the estuaries compared with the cross section of the mouth, however, these estuaries are "choked." The lower well-mixed sections flush rapidly in the larger estuaries, but the upper regions may take two to four times as long to flush. Successive upstream segments of the estuary take longer to flush, and at one point in the upper estuary the water present at high tide cannot reach the estuary mouth and exit during a single ebbing tide. Most of this water from the upper reaches is returned on the succeeding floodtide to the region of its previous position. This is a seasonal null point. Above this point, the water mass is only gradually replaced by streamflow and tidal mixing, so that it attains a characteristically high salinity in summer and an elevated temperature in shallow areas from warming by the sun.
Both floating and suspended woody debris are retained longer in upper estuarine regions with the longest flushing times, whereas floating wood in the lower segments exits more rapidly. Marine wood borers adapted to the annual salinity and temperature regime of the upper reaches grow and reproduce rapidly at the warm temperatures reached above the summer null point. Decreased flushing also increases the chance that the dispersion stages of wood borers will be retained longer in the upper estuary and thus increases their chance of encountering new wood that is also retained longer in the upper estuary.
Velocity of ebb tide current may be high at the mouth of Northwest estuaries (Bourke and others 1971). Maximum ebb currents may be 6 knots per hour at the mouth of the Columbia River, as much as 7 knots at Coos Bay, and 5 knots at the entrance to Grays Harbor-sufficient to move floating wood rapidly away from the immediate coast and into the coastal current system.
A tidally related, clockwise rotating eddy has been identified just off the mouth of the Columbia River (Barnes and others 1971). The eddy is strongest at ebb tide and brings floating material onshore just north of the river mouth. Large rafts of drift trees off the mouth of the Columbia River, noted in the historic record of early explorations, may have been related to this eddy. Such an eddy would have collected trees washed out of the Columbia and retained them in the near-shore ocean.
Page 89
Much of the water leaving the lower estuary on the ebb tide mixes with outside ocean water and is moved away from the mouth by the longshore current (Johnson and Gonor 1982). The water re-entering the lower estuary on the incoming tide is thus mostly oceanic in temperature and salinity. Outside ocean water is thoroughly mixed with water within the estuary during every tidal cycle below the summer null point. The summer and, to a lesser degree, the winter environmental regime of the lower parts of Pacific Northwest estuaries closely resembles that of the cold nearshore ocean. The marine wood borers of this lower estuarine region cannot tolerate upstream conditions in summer, where the water is warmer, less saline, and more characteristic of southern waters. Because of longer water retention times in the upper estuary, the colonization and survival of warm-temperate wood-boring speciesare favored once they are introduced.
Marine, Wood-Degrading Organisms
The fate of trees entering estuarine and marine portions of the ecosystem is mainly determined by organisms that degrade the wood and recycle its nutrients and energy. The kinds of organisms involved and their relative roles in marine and terrestrial environments vary widely. Marine organisms known to degrade wood entering the sea belong to taxonomic groups different than those dominating this process in terrestrial and freshwater environments. They are not as diverse as those in terrestrial environments, and the relative roles of micro-organisms and animals in initial attack on cellulosic materials differ. Marine fungi and bacteria appear to play minor roles in the initial invasion and degradation of wood in the sea. The major roles in degradation of marine wood are played by a few genera of wood-boring crustacea and a larger number of bivalve mollusks (table 4.3). In contrast to the fate of fallen trees in terrestrial environments, wood in estuarine and marine environments is rapidly attacked by animals, long before significant soft rot takes place; the animals disperse it before, not after, microbial decomposition of its constituents.
lsopods
Wood-boring isopod crustacea (gribbles) of the genus Limnoria are major reducers of wood in estuarine and marine waters of the Pacific Northwest. Only two species are involved, L. lignorum, the endemic northern species, and L. tripunctata, the introduced species that is predominantly southern in distribution but is also found in the upper reaches of Pacific Northwest embayments and estuaries. Limnoria lignorum is a coldwater species that occurs in embayments and the lower reaches of estuaries. It can reproduce in the cold outer coastal waters where L. tripunctata can survive as adults but can rarely reproduce or establish new colonies.
Both species are typical isopods in appearance, resembling small, elongate pill bugs of terrestrial environments. Limnoria lignorum is large; individuals average 0.012 to .016 inch long and 0.004 inch wide. Adult L. tripunctata are smaller, typically about 0.012 inch long and 0.004 inch wide. Although both gribble species are small animals, they are major wood-deteriorating organisms in the sea because (1) they live in dense colonies in interconnecting burrows in the surface layers of sound submerged wood, and (2) they are abundant and widely distributed. They do not attack rotted wood, but bore actively and continuously by removing small pieces of fresh wood from the blind ends of their burrows and consuming them. All wood removed by gribbles during burrowing is ingested. About 45 percent by weight is used and the rest is voided as fecal pellets (Ray 1959a, 1959b). These small, flattened, cylindrical pellets are composed of finely ground wood fibers lightly bound together by a thin membrane of chitin secreted by the hind gut. Fecal pellets are continually washed from the gribble burrow systems by respiratory ventilating
Page 90
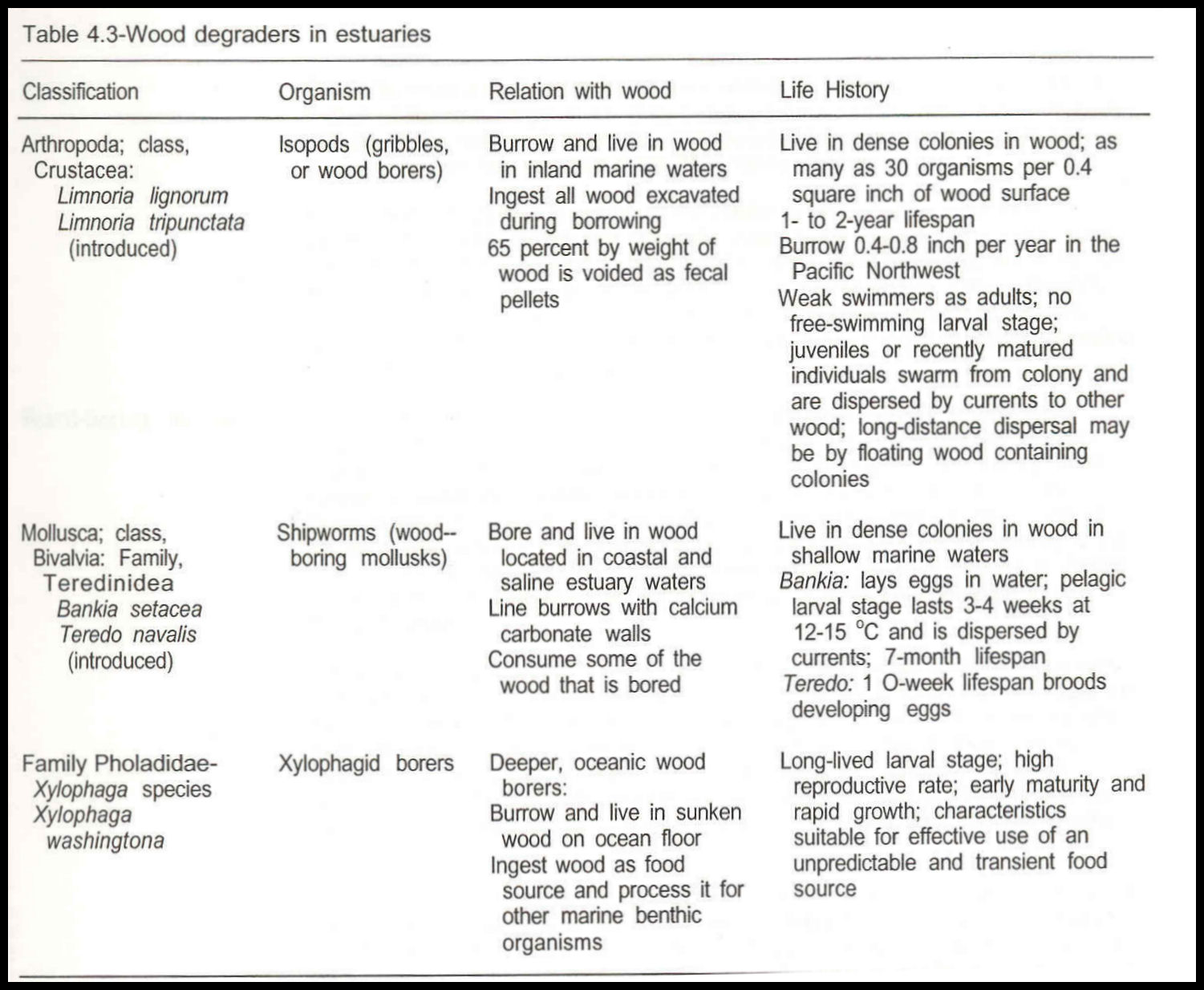
currents created by the animals. The nonbuoyant pellets sink rapidly in still water. They are light enough, however, to be easily dispersed by the typical wind-driven turbulence and currents of estuarine and coastal shallow waters. Gribbles convert wood in one step from the original massive, intact form to microscopic, nonbuoyant particles freely dispersed in the environment and deposited in surficial sediments. As individuals of a colony burrow deeper, the outer, riddled region of the wood with the older galleries becomes waterlogged and softened by micro-organisms. Consequently, it is fragile and easily broken into small pieces by abrasion and wave action that gradually removes the outer layers no longer occupied by the gribble colony.
Gribble population density in wood is high, as many as 30 animals per 0.4 square inch of wood surface. Long-distance dispersion along the coast or between embayments is mainly by colonies in wood that floats from one location to another.
Page 91
Chemical reduction of wood by isopods.-Although gribbles remove a significant fraction of the total weight of wood by passing it through the digestive tract, they do not use all the major polysaccharide components with equal efficiency; and there is no evidence that they can alter lignin (Ray 1959a, 1959b).
Both species of gribbles in waters of the Pacific Northwest are predominantly intertidal or subtidal animals that principally attack wood located in the lower, saline intertidal zone or the portions of large upright pieces, such as pilings, below midtide depth. Gribbles are intolerant of sediment in their burrow system. When colonized logs are rolled by waves across mudflats or sand beaches, the burrow systems become clogged and the colony is reduced or killed. Such wood is readily reinvaded if it is refloated by subsequent tides.
Wood-Boring Mollusks
Other than gribbles, the only shallow-water, wood-boring animals in the Pacific Northwest are two species of shipworms. These are the most important reducers of wood in estuarine and shallow marine waters of the region. The most common species of shipworm is Bankia setacea, a coldwater native to the North Pacific found in the open coastal ocean as well as in embayments and the lower parts of estuaries. Because of its abundance, large size, and rapid growth, B. setacea is the major marine, wood-destroying animal in these waters. The other species is
Teredo navalis, a smaller, introduced warmwater species of restricted distribution in the Pacific Northwest.Shipworms are specialized bivalved mollusks that inhabit shallow water (maximum 650 feet) and coastal areas. They have elongated bodies and use toothed ridges on their specialized shell valves for mechanically rasping their way into wood. As ship worms bore into wood, they secrete a calcium carbonate lining to their burrows. Bankia larvae first attach to wood by a byssal thread and penetrate the wood within 48 hours. Cutting teeth develop on the larval shell after attachment, and the metamorphosed larva begins to bore into the wood, burying itself completely within 24 hours.
These infestations, often so dense that their burrows nearly touch, can rapidly attack and convert the interior of large, solid trees into finely divided wood powder that is dispersed into the estuarine environment. The role of shipworms as agents of wood conversion and dispersion in the carbon and energy cycles of Pacific Northwest coastal waters has not been studied. Bankia sefacea is a highly opportunistic species with rapid growth, a relatively short adult lifespan, a high reproductive output, and widely dispersed, long-lived pelagic larvae capable of rapidly colonizing new wood as it arrives in the coastal waters. In Pacific Northwest coastal waters, infestations of wood by Bankia are intense.
Bankia uses the cellulose fraction of the wood with about the same efficiency as gribbles. Lignins and the remaining cellulosic material are not digested but are voided as finely divided detrital particles in fecal pellets. Shipworms must bore to grow, so much of the bored wood is not ingested but is flushed directly out of the burrow as it is ground into a fine powder. Shipworm fecal pellets are lightly bound with mucus and easily break apart. The fine wood particles they contain are not buoyant but are easily suspended. Shipworm wood borings thus become microdetritus, like those from gribbles, and directly enter the detritus web.
Page 92
Xylophagld Borers
Deep-sea wood-boring bivalves (xylophagid borers) are found off the Pacific Northwest coast. These borers quickly invade wood in the deep sea and grow rapidly. Their boring activity converts the wood into fecal pellets that settle to the sediment surface and attract other deep-sea benthic animals. Their conversion of wood into a readily available detritus source supports the development of a complex benthic community.
Like the shipworms of shallow water, the deep-sea xylophagids are opportunistic species. They quickly colonize wood that, having been carried offshore, becomes waterlogged and sinks to the deep-sea floor. In the North Atlantic, test boards on the floor of the sea (5,948 feet) became infested with a dense population of two species of xylophagids within 104 days (Turner 1977). These deep-sea wood borers have a high reproductive rate, an early maturity, a high population density, and rapid growth-characteristics suitable for effectively using wood that is an unpredictable and transient habitat and food resource in the deep sea. Because adults die as the wood is consumed and disintegrates, dispersion must be through an abundantly produced, efficient, and long-lived larval stage that searches for wood at the sediment-water interface.
Like shipworms, xylophagids use wood for both habitat and food. They are probably the most important deep-sea organisms for converting wood into a finely divided detrital source of food for other deep-sea benthic organisms. They also provide locally abundant prey to deep-sea benthic predators.
Fungi
Marine fungi degrade cellulosic materials, but their ability to rapidly or extensively degrade large pieces of wood appears to be much less than that of terrestrial fungi. Cellulolytic activity has been demonstrated, however, in some taxonomically diverse marine species of Fungi Imperfecti and Ascomycetes. These same organisms have been associated with soft rot attack and weight loss in experimental blocks of wood placed in the sea (Jones and others 1976). Virtually nothing seems to be known about their ecological roles in wood degradation and energy or nitrogen flow in marine detrital systems. Marine fungi are not requisite for the initial attack on wood by the primary marine wood-deteriorating organisms in Pacific Northwest waters. Most of the fungi that break down lignin attack only the wood surface, no deeper than one-eighth inch. Species of the fungal genus Luworthia, however, penetrate deeply into wood and are more like terrestial wood-decaying fungi.
Bacteria
The role of cellulose-degrading bacteria in the sea is obscure, but many species of marine bacteria have been identified as active degraders of cellulose (Kodata 1958). Cellulose-decomposing aerobic bacteria are abundant in seawater and bottom sediments and are widely distributed in the sea and estuaries. These organisms play an important role as remineralizers of cellulose-containing organic matter, but their exact role in marine and estuarine carbon cycles is not known. Some cause rapid deterioration of cotton nets in the sea, and some colonize and attack submerged wood. Cellulolytic bacteria in the digestive tract of the shipworm Teredo navalis are probably responsible for the breakdown of ingested cellulose. Previous or concurrent direct attack on wood by bacteria, however, is not a prerequisite to marine borer activity. Bacteria seem to be important in the breakdown of finely divided wood particles, but the intermediate ecological roles of cellulolytic bacteria in the degradation of cellulosic detritus produced by marine wood borers have not been evaluated.
Page 93
Fallen Trees in the Tidally Influenced River
High water, especially winter storm floods, brings large trees into the tidally influenced region of coastal stream systems. Tides influence water level even in the extreme upper region of the estuary where the salinity is too low for marine wood-boring animals to survive. Surface salinity is virtually zero, but bottom water may contain some salt in summer. The ecological functions of fallen trees in the tidal river water are much the same as those upstream, but many of the fish and other animals are different from those upstream. Fallen trees and logs are present in channels and on intertidal banks, including any freshwater marshes in the reach under tidal but not salt influence (Secretary of the Treasury 1859) (fig. 4.4).
A large drift tree whose roots embrace a mass of hard clay or stones...may very readily be carried outward by the strong effluent current of the Columbia, and especially in the season of freshets; but when the current slacked the root of the tree would remain upon the bottom, sink into the sands, and continue stationary whilst the remaining movement of the current would be able to shift the loose sand and deposit it around the roots.
Fallen trees influence the estuarine portion of the ecosystem, mainly through their physical properties as large masses; they form heavy, solid objects and firm substrates in an environment where the bottom consists mainly of fine sediment. Fallen trees in the tidal river segment of coastal stream systems create riffles and provide shelter from predators for upper reach fishes. Examples of common fishes in this section of Pacific Northwest estuaries are stickelback, sturgeon, starry flounder, and juvenile and adult salmonids. Fallen trees can also affect local waterflow patterns by creating turbulence and thereby affecting the sedimentation pattern and the formation of bars or mudbanks. Emergent parts of fallen trees stranded in the channel or partly or wholly on tidally exposed banks are used by water birds as refuge perches during daily rest cycles, or by predatory birds, such as herons and eagles, as hunting perches.

Figure 4.4-Rootwads help anchor driftwood (photo, courtesy of the Oregon Historical Society, negative ORH 45285).
Page 94
Fallen Trees in the Tidal River
Biological decay in the uppermost estuary (freshwater, but tidally influenced river section) is slow, on the order of many decades, especially for trees partly embedded or buried in typically anaerobic upper estuarine muddy sediments where attack would be only from cellulolytic fungi. Unlike the saline lower estuary, there are no animal groups to act as rapid degraders. Battering by trees rafted downstream on floods or uplifted and carried by tides is probably the major force breaking trees into smaller components. Rolling breaks branches, and trees are typically caught on tideflats by stubs of broken branches.
Fallen Trees in the Upper Estuary
Fallen trees are further transported by floods and tides from the tidal river into the upper estuary system, that brackish, tidally influenced region above the summer null point. Salinity in summer at both surface and bottom can be appreciable but less than in the more marine lower reaches. Salinity in winter is very low, and most marine animals not adapted to prolonged lower salinity cannot survive. Because of lower streamflow and limited tidal flushing in summer, water temperatures in the upper estuary are significantly higher than those in the lower reaches, so marine wood borers are not present.
The gribble Liminoria tripunctata is found in marine waters from inland British Columbia through the Puget Sound region and in all the major estuaries of the Douglas-fir biome. The introduced shipworm Teredo navalis is established on the Pacific coast in San Francisco Bay and in the inner parts of a few embayments on outer Vancouver Island. It is more resistant to periods of lower salinity and higher temperature than the native shipworm Bankia setacea and consequently can establish populations in upper estuarine areas where Bankia cannot colonize. Trees and logs caught in the channels, or stranded, or embedded in the banks and tideflats of the upper estuary are soon attacked by the gribble L. tripunctata.
Attack by gribbles provides continuous and rapid physical breakdown of the wood surface that reduces the radius as much as an inch a year and produces large quantities of fecal pellets. The easily suspended fecal pellets are a carbon source of the sediment surface and the midwater pelagic system. Gribbles transfer fine wood particles to the carbon pool of the benthic sediment system by enormously increasing the surface area of wood and effectively converting trees directly into nonbuoyant wood powder. Further microbial degradation of gribble fecal pellets by cellulolytic bacteria in the surface sediments may channel this carbon source into the benthic detrital food web. The role of wood transferred to benthic and pelagic detrital systems in the form of fecal pellets from marine wood borers has not been examined. Organic detritus is the principal energy source for food webs in estuarine and shallow marine benthic portions of the ecosystem; the principal source of this detrital carbon is debris from macrophytes in the system. Some of the pelagic food web is also based on detritus, and fecal pellets from gribbles are an appropriate size for direct ingestion by zooplankton. Attack by marine borers dramatically increases the rate at which the wood becomes waterlogged and sinks. The role of such wood as large, solid, inplace objects is similar to its role already described for the upper estuary.
Sediment lodges in the extensive spaces of abandoned outer burrow systems of gribble colonies, and some sediment fauna are found in the old, riddled wood. As the surface area of the hardened wood increases, the older portions are invaded by bacteria and marine lignicolus fungi that attack the remaining wood.
Page 95
Recently arrived, buoyant trees and logs in the upper estuary are transported by river floods and winter high tides to the extreme edge of the highest reach of winter high waters, the highest of the year. This usually results in a dramatic strandline of large trees and logs, sometimes in continuous rafts, in the upper or high salt marsh that is the community type characteristically present at this tide level within the upper estuary. Trees and logs function here as agents of disturbance in an otherwise very dense, productive, and uniform plant community, the consequences of which we discuss later (fig. 4.5, color).

Figure 4.5-Driftwood deposited by storms in the high marsh can serve as nurse logs to the forest surrounding the estuary.
Fallen Trees in the Lower Estuary
The lower estuary, from below the null point to the mouth, is dominated by marine influence. Its salinity and temperature range are nearer to those of the open coast than to those of the upper estuary. Trees and logs here are degraded to smaller detritus but have the same physical role as in the upper reaches of the estuary.
Trees and logs transported into the lower region of Pacific Northwest estuaries enter a habitat too cold in summer for reproduction by L. tripunctata. Lower estuarine salinity regimes, however, are sufficient for occupation and survival by the two indigenous species of marine wood borers, L. lignorum and B. setacea. Both can colonize wood for most of the year, and Bankia, in particular, has a high growth rate and can significantly degrade large pieces of wood within 1 year. Between the two, any stationary wood in the lower estuary is rapidly destroyed. The result, again, is reduction of large masses of wood to a fine, nonbuoyant wood powder, most of which is introduced into detritus-based food webs of the estuary. Both the mass and the physical properties of trees are altered; waterlogging increases, and the surface of the trees is converted into a friable mass of riddled wood rapidly abraded into macrodetritus by waves and battering. Shipworm wood borings and fecal pellets become microdetritus and directly enter the detritus web.
Fallen Trees in Salt Marshes
Salt marshes are densely vegetated, low coastal wetlands at elevations within the annual vertical range of regular tidal fluctuations. Plants of the salt marsh community are capable of growing in saturated estuarine sediments and withstanding stresses from salinity and tidal inundation. The requisite conditions of lower salinity, extensive areas of soft sediments at high tide levels, and low wave energy are virtually restricted to estuaries in the Pacific Northwest.
There are no true open coast salt marshes in the Pacific Northwest. Estuarine marshlands have built relatively rapidly toward the sea by deposition of sediments influenced mainly by modern economic activities. Extensive areas have been removed from estuarine systems through diking for agriculture, pastures, roadbuilding, and construction. Logging, road construction, and cultivation-all of which disrupt watershed stability-increase sediment loads in rivers and streams and result in increased sediment accumulation rates in estuaries.
Salt marshes are important parts of estuarine systems in the Pacific Northwest. Salt marshes have very high annual plant production rates, a significant fraction of which is exported to the rest of the estuarine portion of the ecosystem as plant detritus. They also function as hydraulic buffers to flood and storm surges because of their extensive area. Migratory waterfowl and juvenile fishes, especially salmonids, use tidal drainage creeks in the marsh at high tide.
Page 96
Eilers (1975) examined the role of trees in salt marshes of the Nehalem estuary in Oregon through the use of historic charts and aerial photographs and by direct observation. Information indicated that trees have long been important in salt marshes. The upper margins of the high salt marsh were clearly delimited in 1939 aerial photographs by extensive and continuous piles of large drifted trees. Trees, many of them large, were abundant enough to obstruct the channel in the period around 1900. An island, which first appeared in the Nehalem estuary between 1875 and 1914, was covered with live trees and fallen trees associated with other changes in topography. The island today stands 6 feet above tide level, illustrating the sediment trapping ability of piles of down trees and associated vegetation.
Eilers (1975) found that circular-to-oblong depressions, 1.8 to 6.8 feet in diameter and 0.8 to 1.6 feet deep, common in the high marsh at tidal elevations between 9 and 10 feet above mean low water were former resting places of large drifted trees that had been moved about by extreme high tides. When drift trees remain in place in marshes for long periods, the general elevation around them increases from siltation and accumulation of organic matter. When these trees refloat during unusually high tides, floods, or storm surges, the shallow depressions that remain in the marsh increase habitat diversity; at low tide, these depressions are filled with juvenile fishes. Refloated trees are an important source of disturbance that also influences the distribution of marsh plant communities and plant succession (fig. 4.6, color).

Figure 4.6-Sitka spruce and huckleberry on nurse log in Nehalem Bay, Oregon.
The relative mobility of large drift trees and logs in different parts of the high marsh influence successional processes in opposite ways. A spruce/alder/willow wetland forest community dominates the highest portions (9.4 to 9.8 feet mean lower low water) of the salt marsh, slowly invading it as the progradation characteristic of marshes in this region proceeds (Eilers 1975, Johannessen 1964). This forest advances in some places by active colonization of stable piles of drift trees and logs left on the high marsh by past storms. Large drifted trees embedded in the marsh are colonized by terrestrial plant species unable to grow directly on the marsh soil because of salinity. Most of the spruce in the high marsh were growing on nurse trees, and few of the spruce roots extended into the marsh soil (fig. 4.7). In other parts of the marsh exposed to winter storm waves, the forest edge was in retreat, mainly because accumulations of drifted trees battered against the trees and upper marsh shore at abnormally high tides (Eilers 1975).
Drifted trees in the upper marsh are exposed to a wet, predominantly freshwater environment and resemble decaying fallen trees in terrestrial environments rather than trees elsewhere in the estuary. If these trees remain in place, they are attacked by wood-decaying fungi and show the typical sequence of internal rotting. Their relation to nitrogen cycles in the upper marsh and to small mammals inhabiting this environment is not known.
Page 97

Figure 4.7-Sitka spruce grow on nurse logs in Oregon coastal marshes.
Use of Drift Trees in Estuaries by Birds and Seals
Much of the following information about birds’ use of emergent portions of drifted trees and logs in estuaries is from Bayer’s (1978, 1983) extensive studies of birds’ use of estuarine resources in Yaquina Bay, Oregon.
Bald eagles use trees and logs away from shore on tideflats and on sides of estuary channels; they avoid perching directly on soft mud surfaces. Because eagles forage from a central place, use of perches near shore rather than more distant, taller trees conserves the birds’ energy. They perch in places that provide good visibility of open channel water and of tideflats where they watch for opportunities to capture aquatic birds or steal prey caught by gulls and herons in the shallows on the tideflats (Stalmaster and others 1985). Drift trees on flats are more centrally placed in relation to potential food resources for eagles, so flight time to and from a foraging area is less (fig. 4.8).
Green herons, great blue herons, and great egrets are predators that prefer to stand and wait for prey, principally fish, to pass by. They can wade in the intertidal flats only when the water is no more than 8 inches deep. Herons and egrets use drifted trees that remain partly out of water at times of high tide, as well as floating logs, log rafts, and booms as perches for resting and foraging. Great blue herons defend feeding territories on log rafts when they are available, virtually confining their feeding activity to these territories. They conserve energy by using emergent wood near or surrounded by water to combine two necessary daily activities-perching and foraging. Herons thus have more time for foraging than they would hunting for fish when the tide is low enough for them to wade in the shallows. When perches on tideflats or in the channels are not available, herons must alternate between perching in distant trees to rest and wading at low tide to forage, with flights necessary between perching and foraging locations. Herons in upper Yaquina Bay have established territories on log rafts floating along channel margins. These herons can avoid long foraging flights and require smaller feeding territories because the rafts are over deeper water than they could wade in; thus they can also catch larger fish found in deeper water than they could by wading on the flats where smaller fish usually occur.
Page 98

Figure 4.8-Bald eagles choose to perch on mudflat objects, such as driftwood, rather than on the tidal substrate (photo, courtesy of Range Bayer).
Cormorants and brown pelicans also require perches for rest between periods of foraging, and they prefer perches surrounded by water and away from the shoreline-wooden structures, such as piling, and stranded logs and trees. Small shorebirds that feed on tideflats at low tide and require perches for resting at high tide use any available emergent wood.
Natural, old, drifted trees are more abundant in Siletz and Nestucca Bays than in some of the other Oregon estuaries; birds can nest in young trees growing on drifted trees and logs stranded in the intertidal area. Purple martins, which require rotting wood for cavity nesting, use tree butts sticking out of the flats at Siletz Bay. Because rotting trees on land near water have become scarce, suitable stranded trees become important resources for these birds.
Gull-nesting areas within estuaries are uncommon today because of limited suitable nesting habitat. Pilings, human made structures that resemble formerly more abundant stranded trees, are used by gulls for nesting (Bayer 1983).
Natural drifted trees in estuaries and the use of herring spawn by marine birds in Oregon estuaries are indirectly related. Herring that enter estuaries to spawn require solid substrates in the intertidal area for egg (spawn) attachment. In estuaries where the intertidal area and shore are predominantly soft muddy sediments, solid surfaces are limited. Masses of herring spawn are attached to red algae, rockweed (Focus), eelgrass, rock, shells, and pebbles on the flats and on any available wood, such as logs and drift trees, especially branches of trees stranded in estuaries. Using herrings’ preference for finely divided spawning substrate, Pacific Northwest Indians once placed brush and cut branches in the estuaries during the herring spawning season to collect spawn to eat. At present, suitable intertidal spawning substrate is limited in Oregon estuaries, and herring spawn becomes overlaid and crowded on the substrate. Crowding greatly reduces hatching success by creating oxygen deficiencies in the mass of spawn. Masses of excess spawn break free and are
Page 99
rolled across the tideflats by waves to accumulate on beaches. Both this beached spawn and the spawn remaining attached in place are eaten by birds, including nonaquatic species, such as crows. Gulls feeding alone can consume most of the spawn on a beach at low tide. During the herring spawning period in Yaquina Bay, the total resident bird population increases from about 1,000 to 2,000 birds of each species to a total of about 10,000 birds (Bayer 1980). To the extent that suitable intertidal substrate, such as stranded tree branches, limits herrings’ spawning in estuaries, the availability of herring spawn as food to birds associated with estuaries is limited.
Rafted and stranded trees and logs away from the shoreline and undisturbed by people are potentially important to harbor seals, which require haul-out areas for resting out of the water. These seals now haul out on log booms and log rafts in the channels of estuaries. The availability of log booms during the day increases the time that suitable haul-out space is available to harbor seals. Without this type of resource, seals can haul out on shore only at night when people are not present. Natural rafts and drifted trees stranded on tideflats very likely were once also used for haul-out by harbor seals and may have been one of the limiting resource for seal populations in Pacific Northwest estuaries (Bayer 1981, 1985; Calambokidis and others 1978).
Fallen Trees Rafted Out of Estuaries
Aboard a seagoing vessel off the Northwest coast in heavy seas during a storm in November 1852, James Swan (1857) reported:
The next morning we found ourselves about thirty miles to the westward of the Columbia River, from which a huge volume of water was running, carrying in its course great quantities of drift logs, boards, chips, and saw dust, with which the whole water around us was covered.
How much wood was historically transported to the ocean is not known, though it must have been substantial at a time when most of the riparian zones along the river corridors and in the watersheds were dominated by large conifers. Hundreds of millions of board feet of logs and downed trees have come down into Puget Sound in Washington and Georgia Strait in British Columbia from the rivers that drain the Washington Cascade Range and the British Columbia coastal mountains. These large quantities were joined by significant numbers of logs that were separated from rafts in estuarine log storage areas or from towed rafts (fig. 4.9). More than 10 billion board feet of logs are annually stored or travel in estuaries and lower rivers of the Pacific Northwest from Alaska to California, including British Columbia (Sedell and Duval 1985). A 1-percent-escape rate would allow more than 100 million board feet to enter the ocean from this source alone.
More than 2 billion board feet of wood is estimated to be annually transported to the North Pacific ocean, an estimate believed to be conservative because large regional storms were excluded from the calculation, and pieces of wood less than 10 feet long were not considered (Sedell and Hansen, unpubl. data). Two billion board feet per year is a small amount for the entire northeastern Pacific from San Francisco Bay to Nome, Alaska. This is testimony to the retentive nature of rivers and estuaries, as well as to the substantial reduction in volume and size of wood now entering western streams.
Page 100

Figure 4.9-Logs and other escaped wood added to driftwood in estuaries and on beaches (photo, courtesy of Jack’s Photo Service, Coos Bay, Oregon).
Amounts of wood in rivers and estuaries in the past were high compared with today’s supply. In the past, sources of large, long trees were abundant; a large amount of wood was stored on beaches, in estuaries, and in rivers; and there were no dams on the rivers (fig. 4.10). A vast array of organisms have adapted to the use of this material. Some may find substitute habitats, but others-obligate feeders of wood in the ocean environment -cannot.
Today, only 7 of the 17 coastal port authorities in Oregon are directly involved in snag, stump, and debris removal in their estuaries. The sources of drift have been greatly reduced since the 1850’s, and the opportunities for wood to be retained in the lower estuary have also been reduced through diking, marsh filling, and channelization. Pilings create retention sites, but these sites are likely to be cleared of debris.
Ocean Transport of Woody Debris
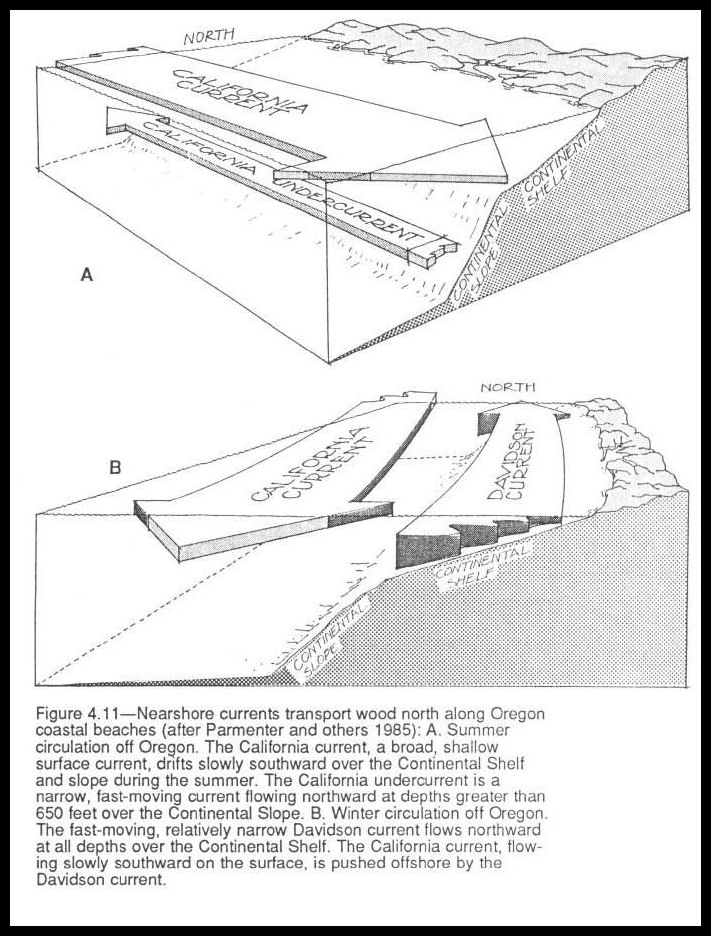
The transport and fate of floating or sunken woody debris discharged from estuaries into the ocean is determined by regimes of coastal wind and current. Along the Oregon and Washington coasts, these regimes differ in direction and intensity with season, so that the direction of wood transported from the same estuary varies greatly. The California current flows south along the coast all year, a broad and slow, surface flow of about 0.2 knot per hour. During the winter, a second coastal flow known as the Davidson current forms and flows northward over the Continental Shelf, pushing the California current somewhat offshore. Its mean velocity is 0.35 knot per hour and results from strong northerly atmospheric circulation.
Prevailing coastal winds shift north-south direction seasonally, with short transitional periods of weak winds. Strong winds blow predominantly out of the southwest from October through April (Barnes and others 1971) (fig. 4.llA). Occasional periods of onshore winds during the summer move a shallow layer of surface water northward along the coast and toward the shore. Storm winds often beach large trees floated out of estuaries and northward along the coast. Redwood stumps from northern California are stranded on Oregon beaches when high tides and storm surges combine to cause unusually high water.
Page 101

Figure 4.10-Beached wood in the late 1800’s often included cut logs as well as drifted trees (photo, Society, negative ORH 26616). courtesy of the Oregon Historical
Page 102
Weaker coastal winds that are more constant in direction blow from the north-northwest from May to September and create a nearshore current that flows south and offshore (fig. 4.11 B). In contrast, when floods bring large quantities of wood downstream into the coastal system, wind and currents keep floating wood inshore and do not allow it to circulate in the open ocean. In summer, offshore movement of water from the nearshore zone is balanced by upwelling of deep water in a zone 5 to 10 miles from the shore. Complex shoreward movements associated with these upwelling fronts may also entrain wood and retain it in the coastal region.
Fallen Trees Along the Shore
Driftwood deposits of the Pacific Northwest differ in both kind and quantity from those observed along other coasts. More important even than the great size of individual trees or logs is the massiveness of the overall deposits, each of which may cover many acres of beach to a depth exceeding 10 feet (fig. 4.72, color). The total accumulation of driftwood between Cape Mendocino in California and Puget Sound in Washington is estimated to be several million cubic yards or a few billion board feet (Stembridge 1979).
Trees are driven ashore on sand beaches by wind and waves along most of the Pacific Northwest coast. These trees form large piles of driftwood, once characteristic of Pacific Northwest shores, until removed by extremely high water. They act as barriers to wind-transported sand and can form the nucleus for at least a temporary accumulation of sand. Drifted trees are often found deep in the base of large dunes and spits when another cycle of waves erodes the sands.
Drifted trees play an important role in natural cycles of erosion and deposition of beaches and foredunes along the Oregon coast. These cycles were examined by use of a series of aerial photographs beginning in 1939 to study long-term changes in the Siletz spit (Komar 1983, Komar and Rea 1976). There, erosional and accretional episodes occur along segments of the shore 325 to 1,950 feet long. The following sequence of events was identified:
1. Rip-currents begin to erode embayments in the foreshore. During periods of high tides and high waves, the deepened embayments permit waves to reach the backshore area and the seaward edge of the foredunes, eroding a vertical scarp into them.
2. Subsequent high tides deposit drifted trees and logs in the embayment at the base of the scarp.
3. Lower energy waves build a broad, high "summer" berm in the embayment.
4. The criss-crossed mass of trees and logs behind the berm effectively trap sand washed in by waves or blown onto the upper beach at low tide by the predominantly onshore winds.
5. Windblown sand continues to accumulate around the trees and logs, reestablishing the foredune, which becomes colonized by dune grass. The foredune often becomes reestablished in 10 to 15 years.
6. Dune reformation is eventually followed by another episode of erosion, repeating the cycle and exposing trees and logs previously embedded in the dunes.
Page 103
More evidence of the influence of driftwood on shoreline stability and accretion comes from a study of the Alsea spit in Oregon (Stembridge 1974, 1979) and from studies of the beaches of northern Puget Sound in Washington (Terich and Milne 1977). Recent changes were charted along Oregon’s 300-mile ocean shoreline, and 110 miles of stable sandy shore and 40 miles of prograding sandy shore were identified (Stembridge 1976). Much of the stability and accretion was attributed to changes in land-use during the 20th century that resulted in an increase in the coastal sediment supply, in the deposition of saw-log driftwood, and in the introduction of European beach grass.
Drifted trees may thus act as passive, stabilizing objects along coastal dune fronts. Because trees shelter or shade some sand, colonizing plants often grow in the moister sand at the base of stranded trees. Other plants, such as Sitka spruce, grow on drifted nurse logs on the backshore. Driftwood not only provides beach stability but contributes moisture and nutrients that appear to be necessary for the establishment of woody vegetation (Stembridge 1979). The role of driftwood in the maritime dune fringe community has not been well studied. Burrows of mice and insects can be found around the highest, oldest, driftwood trees embedded in dunes behind sandy beaches, particularly on shores in a prograding phase.
Trees beached on rocky shores undergo quite different processes. Winter storm waves batter and pile up large quantities of driftwood against the sides of rocky points and headlands. Such trees are highly abraded and roughly splintered, and winter shores abound in the freshly broken fragments of these trees. Most of this splintered and shattered material is partly waterlogged, and much of it is not buoyant. It appears to undergo further direct grinding on rocks into coarse woody debris, some of which lodges among and below rocks, whereas the greater part is washed into deeper water by the waves. Rocky shores are thus another site of direct and rapid degradation of large trees into small fragments capable of entering a detritus-based system through physical rather than biological agents.
Trees have an important ecological role in rocky shore intertidal community structure. Sessile plants and animals in this community compete for the major limiting resource-attachment space. In the absence of other influences, the more successful or dominant competitors gradually exclude others and occupy all surface space, which results in decreased community diversity. Battering by trees at high tide during winter storms is a significant force in opening up patches of unoccupied space, which thereby contributes to the maintenance of a mosaic community of higher diversity (Dayton 1971). Patches of attached organisms or those cleared by tree abrasion produce a mosaic of different phases of colonization and thus prevent complete occupation of all space by the competitive dominants, with concomitant reduction in community species diversity. Disturbances of both physical (waves and trees) and biological (grazing and predation) origin were the major forces that produced structure and order in the space-limited intertidal community of rocky shores (Dayton 1971).
Page 104

Figure 4.13-Driftwood Beach State Park, San Juan Island, shows that some beaches are still (1986) covered with wood.
As much as 50 percent of the trees along the rocky shores of San Juan Island showed saw-cut ends and were obviously from logging operations (fig. 4.13). About 15 percent of the trees had intact root systems, indicating they originated from erosion of shores and streambanks. The remaining 35 percent of trees on these rocky shores were too worn to determine their origin. By contrast, at an open coast site on the Olympic Peninsula, less than 1 percent of drift trees were saw cut, indicating that most of the large accumulations of such wood in this relatively remote location were of natural origin (Dayton 1971). Even today, naturally derived trees play an important role in community processes on some rocky shores. This role is obviously proportional to their abundance and size, which once were greater than they are now.
Present-Day Look at Wood on Coastal Beaches and in Estuaries
Wood volume on coastal beaches varies between locations and fluctuates between seasons. In winter, storms transport wood downriver into the ocean, then deposit it on the beaches. In other seasons, there may be a net loss of wood from the beaches back to the ocean.
Seventy percent of the volume of woody debris longer than 1.6 feet was lost from river-mouth beaches from 1970 to 1984 (Benner and Sedell, in press).
Gone are the mature stands of trees that were the source of large driftwood before the late 1800’s. Wood is being used for fuel in home wood stoves, and the number of Forest Service free-use wood permits (fig. 4.74, color) issued by the Pacific Northwest Region for cutting cords of firewood have increased by more than 800 percent from 1970 to the present (fig. 4.15). Forest practices regulations that directed the cleaning of wood from stream channels during logging operations in the 1970’s in the Northwest also reduced the source of woody material for estuaries and beaches. Fewer logs now escape from lumber operations and other human-related sources (fig. 4.16).
Page 105
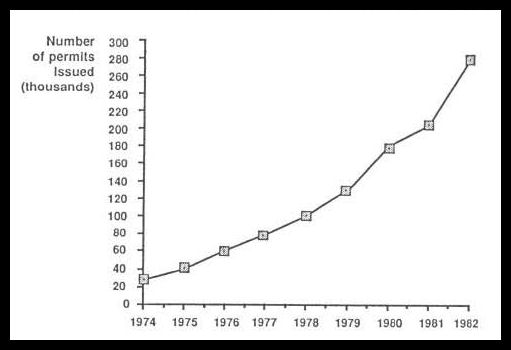
Figure 4.15-Increase in Forest Service free-use permits for firewood, 1974-82.
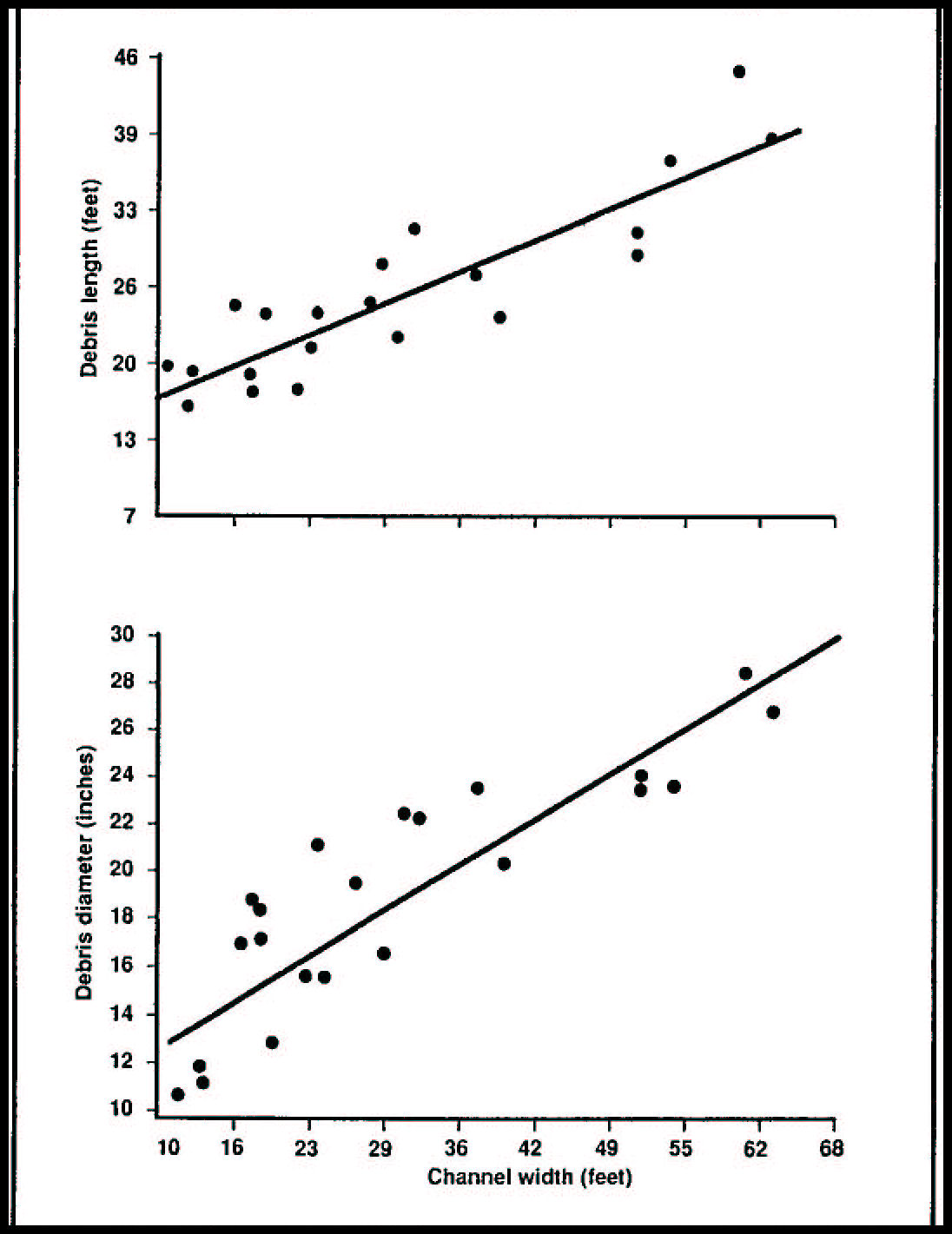
Figure 4.16-Log boom stores logs in an Oregon slough in the late 1800’s. Some logs escaped to the beaches (photo, courtesy of the Oregon Historical Society).
Drifted Trees in the Floating trees can travel long distances and are quickly colonized by shipworms, Open Sea gribbles, and barnacles. When this moving wood sinks to the bottom of the ocean, it forms the primary energy base in a diverse community of invertebrates and functions as an island of productivity in an otherwise stable, low diversity, low productivity environment. Schools of small fish are also attracted to floating objects, including drifting trees and slow-traveling boats. The schools in turn can attract predator fish, such as tuna, and an ecological community becomes associated with the floating forms.
Page 106
Transport and Fates
Drifting trees that escape the inshore tidal currents enter the open sea. In winter, nearshore currents along the Pacific Northwest coast transport trees northward. Unless these trees are moved farther offshore, they will eventually be beached by stormwinds. The direction of transport in the California current system is southerly inshore in summer and offshore all year. If trees escape onshore transport and enter the current, they are carried into the open sea where they may enter the westward transport of the North Pacific gyre.
An inventory of driftwood along some beaches on the Hawaiian Islands showed that most of the trees were from the west coast of North America (Strong and Skolmen 1963). The other trees were native to the Philippines, Japan, or Malaya. Even if attacked by marine borers, large trees remain afloat for long times and, if they are carried into the central gyre system of the North Pacific, may be transported for long distances across the Pacific, to beach in such places as the Hawaiian Islands. Ancient Hawaiian civilizations prized and preferred the huge Douglas-fir trees that washed up on their shores. Local chiefs made double canoes from the large stems, which were described by explorers and missionaries in the early 1800’s (Heyerdahl 1952, Strong and Skolmen 1963). The anthropological records show an integrating of large, floated Douglas-fir and redwood trees into the customs and rituals of Oceania cultures.
Biological Role of Floating Trees
Large trees capable of remaining afloat for long periods of time are natural agents in the dispersion of marine wood borers and attached, sessile plants and animals, as well as of nonswimming mobile animals associated with these encrusting or "fouling" species. Trees at sea are usually heavily laden with growths of such epifauna and flora, including both attached species from the open sea and nearshore species acquired in transit to the open sea. In addition, the water strider, the only insect to have invaded the open sea and become pelagic, lives and lays eggs on driftwood.
Many pelagic fish species of commercial and recreational importance associate with floating objects in the open ocean. Tunas in particular aggregate around trees and logs and other flotsam. Both Japanese and American fishers routinely seek such objects while fishing for skipjack tuna and yellowfin tuna in the eastern and western Pacific (Inone and others 1963, 1968; Kimura 1954; McNeely 1961; Uda 1952), and their success rate for tuna was well over 4:1 in favor of seines set around drifting trees and logs (Pacific Tuna Development Foundation 1979).
Several hypotheses have been suggested to explain this association. One is that shadows cast by floating objects make zooplankton more visible to predators (Damant 1921) or that pelagic fish simply seek shade from the floating material (Suyehiro 1952). Floating trees and logs serve as egg and algal attachment sites, as invertebrate food and habitat, and as cleaning sites where external parasites on pelagic fish are removed (Gooding and Magnuson 1967). Communities of organisms may form food webs in association with drift logs (Brock 1985). Small fish initially accompany a drift tree, feeding on small planktonic organisms in its vicinity. With time, larger predators arrive, taking advantage of accumulated prey. Prey respond by using the floating tree as a shelter. Within 5 weeks from the time of initial object or tree placement, the forms of predators (tunas and dolphin-fish) in the vicinity of the object may vary as much as 100 tons. Predators rapidly deplete the available prey, albeit prey continuously recruit to the object. In their search for food, predators move away from the object to return later, apparently using it as a focal or reference point.
Page 107
Based on underwater and surface observations, dolphin-fish may range from 0.3 to 2.5 miles from a floating tree and return 15 minutes to 1 hour later. Yellowfin and bigeye tuna have been ultrasonically tagged and tracked in Hawaiian waters and found to travel 6 to 12 miles from a floating tree, then to return 10 to 20 hours later (Brill and others 1984).
Size of floating debris has an effect on the attraction of fish to it. Objects with greater dimensions have experimentally been found to attract larger numbers of fish (Hunter and Mitchell 1968). A lower size limit of effectiveness of an object appears to be about 1 square foot. Drifting trees with limbs and roots vertically oriented in the water attract fish better than do trees or logs with horizontal orientation (Yabe and Mori 1950).
Wood in the deep sea not only serves as a source of energy in an energy-scarce environment but also contributes to environmental diversity and associated community complexity in the deep sea. Wood, along with other remnants of land plants, was often present in dredges from the deep-sea floor, even from deep-sea trenches (Bruun 1957). A sunken tree is a focus of abundant deep-sea life for a long time (Bruun 1957, Turner 1977). Wood is more common on the deep-sea floor off the mouths of rivers and wooded coastlines, and deep-sea hauls rich in land-plant debris had a large number and high diversity of deep-sea animals (Bruun 1959; Knudsen 1961, 1970).
Trees escaping into the nearshore system are usually infested with gribbles and shipworms or become invaded while near shore. Such trees gradually become waterlogged and sink slowly if they are not driven ashore by winds. Trees that are attacked by marine borers and sink over the Continental Shelf or in the deep sea represent a direct entry of terrestrially fixed carbon into the oceanic benthic system. The sources, forms, and routes of supply of organic material to the energy-poor deep sea are little known. Woody debris containing deep-sea species of molluscous wood borers of the genus Xylophaga have also been recovered from deep ocean areas. It has been experimentally demonstrated that these deep-sea molluscous wood borers quickly attack wood samples placed on the floor of the sea.
These pelagic teridas have large populations, high reproductive rates, rapid growth, and short lifespans, making them the first known case of an opportunistic species in the deep sea. Like their shallow-water shipworm relatives, they produce large amounts of finely ground wood fragments. Piles of fecal pellets produced by borers may attract more than 40 species of epibenthic, deep-sea invertebrates known to use detrital sources (Turner 1981). The enrichment of the bottom, a result of the disintegration of the wood and the accumulation of fecal pellets, contributes to the development of a rich fauna.
Page 108
Wood also appears to be the normal habitat of some species of small deep-sea mytelid bivalves that attach themselves to crevices in burrows of wood borers. Terrestially derived plant material in the deep sea contributes to niche specialization in groups other than mollusca. Deep-sea species known to be associated with terrestrial plant material include echinoderms that feed on this material, bilaterally symmetrical hermit crabs that live in hollowed plant stems, and gammared amphipods that live in or feed on wood. Chiton, small putilliform gastropods, and sepunculids as well as nematodes and polychete worms are often found on wood or in old burrows of wood borers (Turner 1981). How this wood-dependent deep-sea community finds new wood is still unknown.
Woody debris from decomposition of large pieces of wood on land enters the bedload of streams and is transported downstream to join similar nonbuoyant debris produced in coastal waters and estuaries. This material enters the ocean in large quantities (Hedges and others 1982), to be transported by bottom currents. Near the shore, as far as 1.25 miles from the beach, bottom currents in water less than 130 feet deep result from wave motion and are predominantly a shoreward flow that returns material to the wave zone. Offshore, where water is deeper than 130 feet and thus below the effect of waves, bottom and near-bottom currents on the Continental Shelf flow northward, parallel to the coast.
Vascular plant debris is one-third of the total organic matter in bottom sediments in the Columbia River system (Hedges and others 1982). The major types of plant debris are derived from nonwoody angiosperm tissue and gymnosperm (conifer) wood. The transport of woody plant debris to the sea introduces it into marine food webs as a terrestrially derived organic carbon energy source. The proportions and fluxes of terrestrial plant material into sediments on the Washington continental shelf have been described (Hedges and Mann 1979). The fine sand sediment on the inner continental shelf has a low organic carbon content, mostly derived from marine phytoplankton. On the midshelf, between 12.5 and 25 miles offshore, the sediment is a finer organically enriched silt, as is nearshore sediment around the Columbia River mouth, where the sediment content of terrestrial plant debris would be expected to be high. Ninety-five percent of the organic matter in the sediment off the Columbia mouth is of terrestrial origin, that of nearshore sand is about 20 percent, and the midshelf silts 22 to 25 miles offshore contain 46 to 61 percent terrestrial organic carbon (Hedges and Mann 1979).
Thus, an appreciable quantity of lignin-containing panicles derived from coniferous wood is transported across the Pacific Northwest continental shelf, at least 49 miles offshore, and is deposited in the midshelf sediments where it constitutes at least half the total sediment organic carbon content. The role of intact wood in marine pelagic and benthic food webs remains unknown, as does the potential role of offshore marine wood borers in initial reduction of this wood. The significant amount of coniferous wood products on the floor of the midshelf, however, suggests a potentially valuable research area that could shed light on the interactive link between the forests of the Pacific Northwest and the ocean portion of the ecosystem.
Page 109
Current Management of Wood on Beaches and Estuaries
When logs do reach the shore or beach, should they be removed or allowed to remain (fig. 4.17)? The thinking developed in this chapter argues that abundant drift logs help stabilize beaches by capturing land and shielding the shore from wave action. The States of Oregon and Washington differ in their policy about removing logs. Oregon has the following policy for removal of logs from its beaches:
To assure continuation of scenic and recreational values for public enjoyment at the ocean shore and to protect marine life and intertidal resources, beach logging, as a general practice, shall be prohibited unless such removal can be shown a significant public benefit.
Washington State’s policy on log removal appears to contrast markedly with Oregon's. The Department of Natural Resources (DNR) has an established procedure for licensing individuals who wish to retrieve drift logs, either from the Puget Sound or the Columbia River below Grand Coulee Dam. Certain restrictions are imposed on the license holders, but the department cooperates by administering the sale of unbranded drift or salvage logs retrieved by the licensee. If the log is just floating in the Sound, there is little question about a permit holder’s being able to latch onto it. But if the log is lying on a beach in front of a house, who does it belong to? The DNR's answer is as follows:
Log patrolmen, brand owners, and their agents all have the right to enter peaceably any tideland, marsh, beach, etc., for the purpose of salvaging logs. Likewise, beach owners have rights also; where one right ends and the other begins is hard to define. Generally speaking, the removing of logs from the water side by boat does not infringe upon the rights of the beach owner. A beach owner does not have any legitimate claim to a log that washes upon his beach, and to exercise any appreciable claim over this log could result in criminal action.
But if the beach-front owner believes the logs should be left to protect the beach from wave erosion, the stage is set for argument. The matter is not trivial as more than 9 million board feet were salvaged in 1974 by log patrolmen. The policies of Oregon and Washington apply only to coastal beaches and tidal portions of rivers; neither policy applies to estuaries. The difference is that the beaches in Oregon are public, whereas those in Washington are private.
Page 110
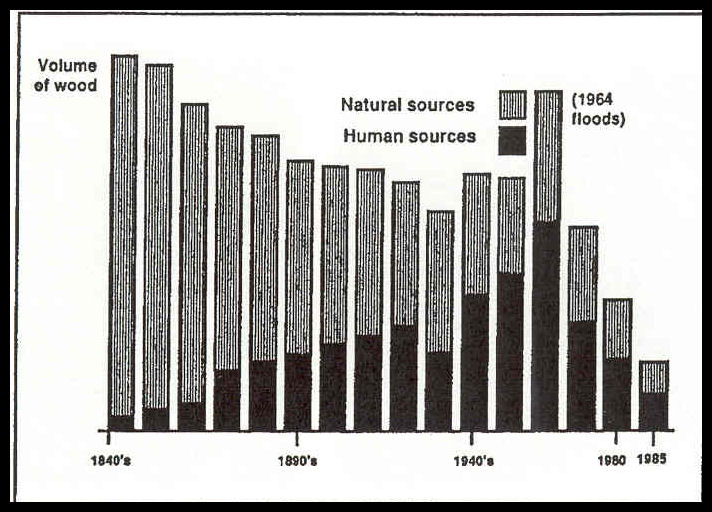
Figure 4.17-Estimates of wood inputs to beaches and estuaries in Oregon from 1840 through 1985.
The other major regulatory arena in estuaries centers on wetlands and mitigation policy and involves the provisions in section 404 of the Clean Water Act of 1972. When wetland or marshes are removed, filled, or dredged, wetlands in another area within the estuary must be created, restored, or enhanced. In Oregon, large woody debris is not currently an issue in the mitigation process. The Division of State Lands and the Department of Environmental Quality, which administer wetland and marsh alteration permits, do not consider woody debris to be an issue in Oregon tidal marshes. They do not encourage removal of wood. There is no mitigation credit for removing wood. Also, in the enhancement and restoration process, wood is not considered. In effect, the Division of State Lands and the Department of Environmental Quality take a neutral stand about wood. They do not encourage removal, and wood is not considered in wetland management. The hope is that the management of the estuary linked to the forest through large woody debris can become a part of the creative mitigation process and can become consistent with the way nature has linked the two.
Page 111
Had color pictures that were placed in the document.
Page 112