
The Unseen World of the Fallen Tree
Decaying trees comprise considerable accumulations of mass and nutrients in unmanaged, old-growth forest ecosystems (Triska and Cromack 1980). Some of the largest accumulations occur in the unmanaged forest stands of the Pacific Northwest. Coarse woody debris can range from 130 to 276 tons per acre (118 to 251 t/ha) in stands from 100 to more than 1,000 years old (Franklin and Waring 1980). Although here we are concerned with Douglas-fir, neither decaying wood nor research data are unique to forests of the Pacific Northwest. McFee and Stone (1966) observed that decaying wood persisted for more than 100 years in New York, and Falinski (1978) noted substantial accumulations of coarse woody debris in an old-growth forest in Poland. Appreciable accumulations of fallen trees also occur in the temperate forests of the Andes of central Chile, in the course of long-term forest succession (Veblen 1979). These observations evidence the long-term continuity of decaying trees as structural components in forest ecosystems.
The decomposing wood of a fallen tree serves as a savings account of nutrients and organic material in the forest soil. Nitrogen, for example, accumulates in decaying trees and branch wood in forests of the Pacific Northwest and other regions (Graham and Cromack 1982, Lambert and others 1980, Lang and Forman 1978, MacMillan 1981, Sollins 1982, Swift 1977b). In old-growth Douglas-fir forests, about as much nitrogen accumulates in decaying, fallen trees as in the forest floor. Other nutrients, such as calcium and magnesium, also accumulate in decomposing woody substrates (Cromack and others 1979, Grier 1978, Rennerfelt and Tamm 1962, Sollins and McCorison 1981, Sollins and others 1980).
Nitrogen fixation by free-living bacteria is another mechanism of nitrogen accrual (Cornaby and Waide 1973, Larsen and others 1978, Sharp 1975, Sharp and Millbank 1973, Silvester and others 1982, Triska and others 1982). Although nitrogen fixation in wood is modest compared with that occurring in other substrates in forests, the persistence of decaying wood allows small increments of nitrogen to accrue over many decades.
Further, decomposing wood undergoes changes in other chemical constituents and pH, as well as in physical structure (Aho 1976, Aho and others 1979, Johannson and Theander 1974, Shortle and Cowling 1978, Tatter and others 1971). Very old, decayed wood can even become somewhat humified and leave a long-lasting substrate resistant to further decay.
Fallen trees harbor a myriad of organisms, from bacteria and actinomycetes to higher fungi. Of these, only some of the fungi might be noticed by the casual observer as mushrooms or bracket fungi (fig. 16). These structures, however, are merely the fruiting bodies produced by mold colonies within the log. Many fungi fruit within the fallen tree, so they are seen only when the tree is torn apart (fig. 17). Even when a fallen tree is torn apart, only a fraction of the fungi present are noticed because the fruiting bodies of most appear only for a small portion of the year. The smaller organisms, not visible to the unaided eye, are still important components of the system.
Page 16
Figure 16.-Mushrooms of Inocybe Ianuginosa (Bull.:Fr.) Kumm, a mycorrhizal fungus that fruits only with roots growing in rotten wood.

|_0.5mm_|est.
Figure 17.-Ceratiomyxa fruticulosa (mill) Macbr., a tiny fungus that typically fruits in cracks within brown cubical rotted wood.
Microhabitats and inhabitants
Fallen trees offer multitudes of both external and internal habitats that change and yet persist through the decades. One needs an understanding of the synergistic effects of constant small changes within a persistent large structure to appreciate the dynamics of a fallen tree and its function in the forest ecosystem.
Within fallen trees.-The most obvious changes in fallen trees over time are described in the classification system shown in table 3. The class I tree, mostly intact and undecayed, offers habitat and substrate to only a few primary invaders: bark or wood-boring insects, a few fungi, and bacteria. The entry of these organisms opens the tree to profound changes that render it amenable to many additional tenants. The decayed sapwood enclosed by intact bark in the class II tree provides a soft, fragmented, moist environment much of the year. Many organisms inhabiting the class II tree lose out as decay progresses to class III, when the bark and decayed sapwood slough off. Meanwhile, the heartwood becomes increasingly decayed, but the process is much slower than in the sapwood so the class IV stage lasts a long time.
It is in the class IV stage that the fallen tree presents the most diversified habitat and hence supports the greatest array of inhabitants. The decayed heartwood is relatively stable, so plants that become established on it have time to grow substantial root systems. Decayed heartwood splits into chunks; roots grow down the resulting cracks as well as along insect channels. Invertebrates-from minute mites to centipedes, millipedes, slugs, and snails-find shelter in these openings and passage along them (fig. 18). Vertebrates, such as salamanders, shrews, shrew-moles, and voles, find cover under debris of sloughed bark and rotten wood alongside the class IV tree; they also find the rotten wood on the underside of the tree crumbly enough for digging tunnels or burrows (fig. 19). Fungi and other micro-organisms abound on the wood itself as well as on the new substrates offered by the feces of animals.
Gradually, the class IV tree breaks down into increasingly smaller crumbles of rotten wood to become class V. The intricate labyrinth of cracks, tunnels, and chambers collapses into a mulchlike mound that is excellent rooting material (fig. 20). It is, however, too homogeneous and unstructured to provide good animal habitat. Because it is now composed primarily of residual materials highly resistant to decay, it does not support as diverse a community of micro-organisms as it did in the class IV stage.
Page 17
Figure 18.-A millipede (Harpaphe haydeniana) in a molting chamber made of organic debris under the loose bark of class III fallen Douglas-fir tree.

Figure 19.-Clouded salamander, an inhabitant of class III fallen Douglas-fir trees in forest openings (photo courtesy of Greg Courtney).

Figure 20.-Two decay class V fallen Douglas-firs in the foreground are decomposed into mounds covered with highlighted Oregon oxalis; the background fallen tree is in decay class IV.
Ultimately, the entire tree is incorporated into the soil. It has gone full circle, having been formed as a product of soil and of photosynthetically captured carbon, it now returns to the soil through release of carbon by decomposition.
Checklists of plants and animals.-There are few checklists of either plants or animals that inhabit fallen Douglas-fir in the Pacific Northwest. No checklist of the micro-organisms in fallen trees of western old-growth forests is available; the subject has hardly been studied. (Higher fungi have been cataloged for many kinds of rotten wood in Europe: Kreisel (1961), Pirk and Tuxen (1957), Ricek (1967, 1968), Runge (1975).) Lawton (1971) listed the mosses that occur on rotten wood or stumps in the Pacific Northwest. Deyrup (1975,1976) has done a thorough job with the insects and has identified about 300 species associated with fallen Douglas-fir. The only published checklist for vertebrates that use fallen trees is for northeastern Oregon (Maser and others 1979).
Page 18
Nutrient Cycling
As carryover structural components of old-growth forests, large snags and large fallen trees contribute to the internal cycling of nutrients. Such snags and fallen trees also interact with hydrological cycles and serve as habitat for numerous micro-organisms and higher plants and animals. For example, in one portion of an old-growth Sitka spruce forest in the Olympic National Park, fallen trees covered about 12 percent of the forest floor. A similar proportion of surface area interactions occurs in many old-growth Douglas-fir forests, which resemble some stands in the Olympic National Park (Christensen 1977, Elton 1966, Franklin and Waring 1980, Franklin and others 1981, Grier 1978, Triska and Cromack 1980). Fallen trees in various stages of decay covered 21 percent of the floor of an old-growth Douglas-fir stand in the Cascade Range in western Oregon (Fogel and others 1973).
Fallen trees interact with nutrient cycling processes in a forest through such mechanisms as litter fall (freshly fallen or slightly decomposed plant material from the canopy), throughfall (rain or dew that picks up nutrients as it falls through the canopy), nitrogen fixation, and nutrient uptake by plants associated with the fallen trees. Litter fall and throughfall are major pathways for the flow of nutrients and energy within forests (Carlisle and others 1966, Rodin and Bazilevich 1967, Sollins and others 1980); they contribute nutrients and water to rotten wood. The larger a fallen tree, the more litter it accumulates on its surface and the more nutrient-rich moisture it intercepts from the canopy. The moisture gathers nutrients as it passes through the accumulated litter and soaks into the fallen tree. In addition, a snag may accumulate moisture-carried nutrients and have a higher nutrient capital when it falls than does a live tree.
Ground contact by fallen trees creates opportunities for various interactions with the biotic components of soil and litter. Fungi, for instance, translocate nutrients within the soil-litter system, as both decomposers and root symbionts. Fungi also immobilize translocated nutrients and thereby enrich the decomposing wood substrates they inhabit. In addition, the colonization of decomposing fallen trees by nitrogen-fixing bacteria permits additional nitrogen accretion within the decaying wood (Cornaby and Waide 1973; Dowding 1976, 1981; Larsen and others 1978; Melin and Nilsson 1950, 1952; Silvester and others 1982; Sollins and others 1981; Swift 1977a 1977b).
Colonization of decomposing wood by animals helps microbes to enter interior surfaces of the wood and creates additional openings for entry of water and nutrients; and penetration of the wood by roots of trees, such as western hemlock, facilitates entry by mycorrhizal fungi (fig. 21). Western hemlocks colonize rotten wood over many decades (figs. 9,10, and 14) to insure long-term interactions by root zone processes. Decaying wood thus serves as a savings account of soil organic materials and nutrients in forest ecosystems (Christy and others 1982, Graham and Cromack 1982, Kuhnelt 1961, Lambert and others 1980, Swift and others 1979).
Interactions-Nutrients, Plants, and Animals
The continuum of a fallen tree is composed of, and driven by, an increasingly complex network of simultaneously developing minisystems-all interdependent. These minisystems are: (1) animal-plant-nutrient, (2) plant-nutrient, (3) plant-plant, (4) animal-plant, (5) animal-animal, and (6) nutrient-plant-animal-nutrient. The cumulative effect of these systems is far greater than the sum of their parts. Ausmus (1977) stated the impact simply: ". . . wood decomposition represents a long-term stabilizing force within the forest ecosystem."
Page 19
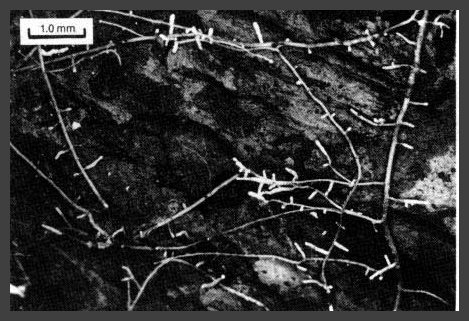
Figure 21.-Small roots and mycorrhizal feeder root tips of western hemlock in rotten wood.
The following simplified scenarios illustrate known facets of each minisystem, and each internal minisystem builds on the preceding one. They are continually being added to, subtracted from, and variously interwoven (Kimmey and Furniss 1943). Some grossly simplified activity levels might be:
1. Animals that eat the fallen tree, which opens it to the outside and initiates. nutrient cycling: wood-boring beetles, carpenter ants, termites, and wood-tunneling mites.
2. Micro-organisms that live on the fallen tree: bacteria, yeasts, and ambrosia fungi.
3. Animals that eat live micro-organisms that grow on the fallen tree: collembolans, mites, and ambrosia beetles.
4. Live plants, such as western hemlock or huckleberry, that form a specific symbiotic (mycorrhizal) association with other live plants (particular fungi), both of which are partially or completely rooted in the fallen tree.
5. Animals that depend on the fungal portion of the mycorrhizal association for their food supply: mites, beetles, and the California red-backed vole.
6. Animals that eat live animals: mites, spiders, pseudoscorpions, centipedes, and salamanders.
7. Animals that eat detritus (dead plant and animal material and animal feces): earthworms, mites, millipedes, isopods, and earwigs.
Trees fall on the forest floor in various stages of decomposition, from sound windthrown trees to snags to stubs of old snags toppled because they are so badly decayed that they can no longer support themselves. Each is unique when it begins its new "life" as a fallen tree. Some of the decay organisms living in standing trees in an old-growth forest may continue their activity after a tree falls. Others, especially those that were in the tops of the standing trees, soon die out after the tree falls and are replaced by ground inhabitants. In the following discussions, we describe the events and organisms in live trees that were windthrown. Fallen snags or stubs are recruited at more advanced stages of decay.
Animal-plant-nutrient.-Tree bark is such an effective protective barrier that the first minisystem to develop in a new, class I fallen tree is the animal-plant-nutrient minisystem. For example, a bark beetle (Scolytidae) (an animal) chews through the bark and thus connects the outside world with the inside of the tree. As the beetle enters and begins to use the tree, it not only introduces fungal spores but also initiates the nutrient cycle with its first deposit of bodily waste.

Page 20
The character of the available food is decidedly varied in the different parts of a newly fallen tree. Proteins are concentrated in the living tissues (phloem-the inner bark and cambium); carbohydrates, on the other hand, are concentrated in the dead woody tissue (xylem). In addition, the living inner bark and cambium are more easily digested than is the sapwood; but the moist sapwood is more digestible than the drier heartwood. So each portion of a fallen tree supports a characteristic group of insects adapted to a specific set of circumstances. Further, the numbers of a given species are regulated by the availability (quantity and quality) of their food supply. Since the inner bark and cambium furnish the most nutritious food, this microhabitat is promptly occupied. The area of next importance is the sapwood, then the heartwood, and finally the bark (Anderson 1960, Graham 1925).
The insects occupying the live inner bark and cambium must be able to chew or bore their way through the dead outer bark and are called borers. There are two groups: (1) those that spend all their developmental or immature stages in this substrate and (2) those that spend only part of their developmental stages in this substrate (Anderson 1960, Graham 1925).

The first group is typified by a bark beetle. For example, the Douglas-fir beetle breeds in both live trees and class I fallen trees that result from windthrow. Because these beetles depend on the fresh, green tissues of the inner bark and cambium, they must develop rapidly before their perishable habitat and food supply dry out or become chemically and physically altered by other organisms.
A female Douglas-fir beetle attacks class I wood in early spring by chewing through the outer bark. When she reaches the inner bark and cambium, she chews an egg gallery that is usually 2feet (60 cm) or more in length along the axis of the tree. The egg gallery is about one-quarter of an inch (6 mm) wide and has small "grooves" on alternate sides; 10 to 36 eggs are laid together in the grooves. When the eggs hatch, the larvae chew lateral feeding galleries through their food supply-the phloem. As the larvae grow, the feeding galleries increase in size. Larvae pack their feeding galleries with borings (refuse) and frass as they eat. The larvae dine throughout the spring, summer, and autumn. In autumn, a larva creates a pupal cell at the end of its feeding gallery where it over-winters as a mature larva. Adults overwinter also and begin to emerge in April. The cycle, from egg to sexually mature adult, requires about 1 year, and one generation is produced annually (Anderson 1960, Chamberlin 1949, Furniss and Carolin 1977, Furniss and Orr 1970, Graham and Knight 1965).

The second group is typified by some members of such wood-boring beetle families as Buprestidae (flatheaded or metallic wood borers) and Cerambycidae (roundheaded wood borers). Some flatheaded wood borers, for example, develop in the inner bark and cambium, but others feed there only for a time and then enter the sapwood. Still others go directly into sapwood and heartwood.
Page 21

Figure 22.-The golden buprestid prefers the sapwood and heartwood of class I fallen Doug-fir trees.
A good example of the family Buprestidae is the golden buprestid (fig. 22). The golden buprestid feeds briefly in the cambium but prefers the sapwood and heartwoods of class I wood. It will, however, inhabit partially to completely seasoned wood.
A female deposits her eggs in flat masses wedged in crevices in the bark or in cracks in exposed wood. The young hatch and immediately start boring into the wood. Their oval tunnels increase in size as the larvae grow. The tunnels range in length from 3 to 15 feet (0.91 to 4.6 m) from where the eggs hatch to where the larvae mature. As they feed and grow, the larvae pack their tunnels with borings and frass. The larvae mature in 2 or more years, construct pupal cells near the surface of the fallen tree, overwinter, and emerge in spring as adults. Before laying eggs, the newly emerged adults feed on needles of Douglas-fir, then find a class I fallen tree and start the cycle again.
Although the normal life cycle of the golden buprestid from egg to mature adult is usually 2 to 4 years, the length of the cycle is influenced by the quality of a larva’s habitat. In poor quality habitat, the life cycle may take a decade or two (Anderson 1960, Chamberlin 1949, Furniss and Carolin 1977).
Within the family Scolytidae is a group of bark beetles called ambrosia beetles that live primarily in the sapwood of class I trees. They are unique among the wood-boring insects in that they do not eat the wood, so the borings are cast out of the tunnels where they collect on the surface of the bark or wood as light-colored powder. Adult beetles, depending on the species, construct a variety of tunnels as follows: (1) an open cavity; (2) a long, winding, branched or unbranched cylindrical gallery in which larvae move about freely; and (3) a compound tunnel, in which "larval cradles" or small pockets are chewed, at right angles, along the main channel (Barras and Perry 1975, Francke- Grosmann 1967, Furniss and Carolin 1977). Ambrosia beetles, especially the females, store certain fungi in specialized structures called mycangia. The fungi, called ambrosia fungi, are introduced into the beetles’ galleries during the beetles’ burrowing. Particular species of fungi are host specific to certain species of beetles. As the fungi grow, they are eaten by both the adults and the larvae. The beetles’ requirements, however, are very exacting; if moisture conditions in the galleries are unsuitable, the crop of fungi either fails and the beetles starve or the crop explodes and the beetles smother in their own food. Hence, the stage of decomposition of a fallen tree is critical (Furniss and Carolin 1977).
Page 22

Ponderous borer
The ponderous borer (family Cerambycidae), the largest western cerambycid, directly penetrates the sapwood and heartwood. Adults emerge during the summer. After they mate, a female deposits her eggs in crevices in the bark or in cracks in exposed wood of class I fallen trees. The eggs hatch and the larvae chew their way into the sapwood and then deep into the heartwood of the tree. They grow to nearly 3 inches (7.5 cm) in length by the time they are mature; one generation requires 3 to 7 years to mature. The oval, meandering galleries of mature larvae are large (1 to 2 inches (2.5-5.0 cm) in cross section) and are filled with frass and refuse. When they are mature, larvae construct pupal chambers at the ends of their feeding tunnels and pupate. They emerge as adult beetles in summer, mate, lay their eggs, and die (Anderson 1960, Chamberlin 1949, Furniss and Carolin 1977).
Another penetrator of fallen trees is the carpenter ant. The abundance of dead wood in an old-growth forest provides numerous sites for queen carpenter ants to establish new colonies, but as these sites deteriorate, the queens die, and the weaker colonies subsequently become decadent and also die. Although fallen trees, stumps, and the bases of snags serve as initial nesting sites, they are not permanent. "Permanent" nesting sites are provided by living trees. The reproductive success of carpenter ant colonies is related to the age of the colonies, and large (old-growth) trees provide permanent nesting sites. When a nesting site finally deteriorates, a strong ant colony can move to another site and withstand catastrophes that would eliminate a weak colony (Sanders 1970).
In brief, the carpenter ant’s life history is as follows: the young males and females leave the nest in early spring and, being winged at that time, disperse in all directions. The air is sometimes filled with these flying ants that emerge simultaneously from many nests in their mating flight (Graham and Knight 1965).
The males die shortly after mating, but the young, mated females may go into old, established colonies to replace decrepit queens or each may establish a new colony. In the latter case, a young queen seeks a small cavity (for example, in a fallen tree) and constructs her brood cell by completely enclosing the cavity, leaving neither exit nor entrance. In this sense, members of the genus Camponotus differ from ants of other genera in that a queen works alone in founding a colony (Graham and Knight 1965).
Once the brood cell is complete, the young queen breaks off her wings; they are no longer of use. And after sealing herself in, she does not feed again until her first brood of young is mature. She lays a few eggs that hatch in about 10 days. The newly hatched larvae are fed a secretion from the queen’s salivary glands. The larvae complete their development, spin their cocoons, pupate, and emerge as adults about 30 days after hatching. The only food the first brood has during their development is the material from the queen’s salivary glands. Although this first brood of workers is small, they take over the work of the nest as soon as they mature (Furniss and Carolin 1977, Graham and Knight 1965).
The workers cut approximately parallel, concentric galleries that run longitudinally through the wood, primarily where it is soft from decay. These galleries are continually increased to accommodate the enlarging colony. The wood is not eaten, as it is by beetles, but is cast out through openings cut to the outside, sometimes called windows. Excavation of the galleries causes piles of wood fiber or sawdust to accumulate below the access holes. Food secured by the workers is also brought into the colony though these openings (Furniss and Carolin 1977, Graham and Knight 1965).
Page 23
The workers feed the queen, care for the eggs she lays and for the larvae that hatch from the eggs, and feed the larvae secretions from their mouths. The "nurses" continually move both larvae and pupae to the most favorable places within a colony. When young adults are fully developed, the nurses assist them from their cocoons and treat them with the "greatest consideration" (Graham and Knight 1965).
At home in shady, wet places, carpenter ants are catholic in diet. Their menu includes animal food, such as caterpillars of certain butterflies, and sweets, such as honeydew secreted by aphids. Carpenter ants have even been known to shelter aphid eggs in their nests during winter and to carry them out in the spring and place them on plants to develop (Furniss and Carolin 1977). With this exchange of materials, the ants establish a purposeful, physical link between the inside and the outside of a fallen tree.

By the time a fallen tree reaches late class II, it has a relatively high moisture content, but the wood is still sound. At this stage of decomposition, it is suitable for habitation by the Pacific dampwood termite, which sooner or later colonizes most large fallen trees in the coastal forest (Furniss and Carolin 1977).
Colonies of Pacific dampwood termites contain reproductively active individuals (primary reproductives), soldiers (that are sterile), and nymphs; but they do not have workers. The winged, reproductively active individuals, the most familiar form, sally forth on warm evenings in August, September, and October to mate and form new colonies. Tevis (1953) aptly described the termites’ appearance: ". . . on a particular quiet, sultry afternoon. . . . shortly before sundown the forest became alive with the wing-rattling of termites streaming from slit-like holes in damp logs and taking to flight."
Typically, colonies are formed by swarms of primary reproductives. Such individuals appear in established colonies and accumulate for a short period before departing on their nuptial, colonizing flights. During the predeparture period, the primary reproductives are sexually immature. Their sexual maturity culminates as they take wing, in about equal numbers of males and females. Termites are weak fliers and seldom travel far without the assistance of a breeze. They are also vulnerable to predators, such as bats. The survivors, on completing their flights, cast their wings. When a survivor encounters an individual of the opposite sex that has also cast its wings, the pair walks in tandem-the male following the female-as they seek a fallen tree in which to excavate a small nuptial chamber and copulate (Imms 1960, Whitaker and others 1977).
Individuals produced during the early stages of colony development are sterile; winged, reproductive individuals develop when a colony is about 4 years old and contains 400 to 500 individuals. Individual termites are long lived (several years), an important factor in permitting a close relationship between successive generations and an important element in the termite’s social organization (Imms 1960).
Pacific dampwood termites feed on the rotting wood in which they live. They even appear to be attracted to a specific fallen tree when it is ready for their habitation; its readiness depends on the presence of certain acids and aldehydes produced by particular tree-inhabiting fungi. These substances initially attract termites to a fallen tree and then attract them within the tree to the wood that is appropriate for consumption (Becker 1971).
Page 24
A termite can digest the wood it eats because of a mutually beneficial, three-way relationship between the individual termite, cellulose-digesting Protozoa, and nitrogen-fixing bacteria that live in its gut. The Protozoa digest the cellulose in the wood and convert it to a form that the termite can use as food. In turn, the termite’s body provides an anaerobic chamber that is replete with food for the Protozoa. Wood particles eaten by the termite pass into the chamber and are engulfed by the Protozoa that ferment the cellulose. Major products of fermentation are carbon dioxide, hydrogen, and acetic acid. The acetic acid is absorbed through the wall of the termite’s hindgut and is oxidized as energy (Breznak 1975).
Nitrogen is important in the termite’s diet, especially because it is required by the cellulose-digesting Protozoa. Wood, the primary ingredient of the diet, is low in nitrogen, although termites consume mostly wood colonized by fungi that probably supply vitamins and some nitrogen (Esenther and Kirk 1974). Whenever the nitrogen intake by a termite is deficient, the nitrogen-fixing bacteria in a termite’s gut make up the difference. The nitrogen-fixing activity varies inversely with the amount of nitrogen contained in a termite’s diet. The nitrogen-fixing response of the bacteria to changes in dietary nitrogen is rapid. The nitrogen-fixing system is efficient and is potentially capable of quickly supplying a termite with usable nitrogen should its dietary level suddenly drop (Breznak 1975).
The Pacific dampwood termite lives in a series of galleries, eaten in the wood, without any external sign of a colony. Even so, by the time a colony has run its course, it has greatly altered the interior microhabitats of a fallen tree. The abandoned galleries form a major network of passages used by other animals and through which plant roots find easy access and ready nutrition.
Some mites also eat the well-decomposed class IV wood (Hartenstein 1962, Luxton 1972). A particular group of organisms in fallen trees, immature wood-tunneling mites, preferentially feeds on the fecal material of such organisms as wood borers. These immature mites probably consume wood that has already been chewed and partially digested until they develop an adult’s capability of chewing and digesting wood first-hand and can excavate their own tunnels (Wallwork 1967, 1976).
Plant-nutrient.-The succession of plants on fallen trees is mediated by changes in nutrient availability and physical properties over time. Three broad phases can be defined: initial, optimal, and final (Kreisel 1961; Ricek 1967, 1968). Early invaders prepare the tree for later colonization by altering its physical and chemical properties during the initial phase. The altered tree provides the best substrate for a wide array of organisms during the optimal phase. Ultimately, the depletion of nutrients and physical deterioration of the wood during the optimal phase diminish its value for many organisms, so fewer species inhabit the final phase.
Page 25
The initial phase encompasses class I and early class II fallen trees, those with bark intact and heartwood still sound. The earliest use of a newly recruited tree is likely to be by heart rot [see web masters note at beginning on heartrot] or butt rot fungi and associated micro-organisms that inhabited it before it fell. Other organisms quickly enter where its interior is exposed at breaks or splits in the bark. When beetles chew through the bark, they create additional ports of entry. The beetles also carry spores of decomposition fungi that thrive in the beetle galleries, which are excellent incubators filled with nourishment for the fungus and protected by the bark from drying and from temperature extremes. These earliest invaders are opportunistic scavengers-fungi and bacteria that join the insects in exploiting the readily available carbohydrates of cambium, phloem, and sapwood of a recently fallen trunk. As the most easily extracted nutrients are depleted, these early invaders sporulate. The spores are then carried off by the emerging new generation of insects reared from eggs laid after the tree fell.
Meanwhile, free-living, nitrogen-fixing bacteria may be active at low levels in the wood to add to the nitrogen pool available for growth of the wood decay fungi (Aho and others 1974, Larsen and others 1978, Silvester and others 1982). Other nitrogen is added by rain falling through the old-growth canopy, which is rich in lichens that fix nitrogen (Sollins and others 1980); and much of the nitrogen extracted by initial decomposers is still present in their dead cells or in frass, ready to be recycled.
Later in this initial successional phase, the early scavengers are succeeded by fungi more competent in decomposing cellulose and lignin in the sapwood This process moves relatively fast, as long as the bark hangs together on the fallen tree to preserve the incubator environment within. Furrows in the bark on the upper side fill with leaf litter and provide sites for several years for seeds to germinate. Where the bark is intact, seedlings generally die during summer drought. If a seedling’s roots find a crack or hole in the bark and grow into the decomposed layer between bark and wood, however, it may find enough moisture to survive the summer. Western hemlock, huckleberry, and salal commonly become established this way on fallen trees, as do spruces (Christy and others 1982, Eichrodt 1969, Gobl 1968, Minore 1972, Mork 1927). Seedling top growth is generally poor at this successional phase because nutrients are limiting and the roots may not have contacted the mycorrhizal fungi needed for nutrient acquisition (see "Plant-plant"). Root growth, however, may be extensive but it is trapped in the zone of decomposition between bark and heartwood.
A transition from the initial stage to the optimal stage of plant succession on a fallen tree occurs in late class II and class III stages; sloughing of bark is a major event in this transition. When the bark sloughs off, it may take with it some of the plants that have grown through it to form roots in the decaying sapwood Plants that remain will have much of their root systems exposed to air and sun. As sapwood deteriorates and sloughs off, additional plants are removed with it.
As the tree enters class IV (most bark and sapwood removed), it also enters the optimal stage of plant succession. The heartwood is much decayed, usually as brown cubical rot. Wood thus rotted becomes spongy and tends to separate into angular chunks, and the intervening cracks provide interior surfaces for the fruiting of fungi and for the growth of roots. Insect tunnels provide additional passages for root growth. Mineral content of the fallen tree at this stage may exceed the original content because minerals have been added by litter fall from the canopy and by throughfall of rain, have been brought
Page 26
in by animals, or have been translocated from underlying soil by fungi or roots. Nitrogen may be added by similar means and by biological fixation (Larsen and others 1978, 1982; Silvester and others 1982). These circumstances provide an excellent rooting medium for plants. A great variety of fungi, both decomposers and symbionts, thrive in the complex of niches within the fallen tree. Lichens, mosses, and liverworts become established on the surface and stabilize it after the sapwood sloughs off. Hemlocks and other plants become established on the upper surface of class IV trees, but their roots grow through it into underlying soil as well as along its length. Nutrients may be acquired from the tree itself or from the soil.
The optimal stage may last for many decades, but in time the fallen tree enters the final, more depauperate stage of class V. It loses structure and shape and becomes increasingly low and covered with forest floor material that buries the lichens, mosses, and liverworts. Roots of overstory trees and of trees that became established in earlier stages now permeate it and bind it together. New plants seldom become established because of the humus cover and the intense competition of roots already there. Relatively few decomposers remain because only the lignins, the most resistant to decay, are left. Mycorrhizal fungi seem to predominate and are the primary fruiters. The moisture content of such material remains high compared with that of soil during summer drought, and roots and fungi can grow actively in it long after most roots in nearby, dry soil have become dormant or desiccated (Harvey and others 1978, 1979a, 1979b; Place 1950).
Plant-plant.- The earliest interactions between plants in the newly recruited, class I fallen tree are between micro-organisms, the initially invading scavengers. Competition for the readily available carbon, nitrogen, and minerals is simple: The first to arrive gets the most. More subtle interactions may also develop between specific organisms; for example, some fungi produce antibiotics that suppress certain bacteria, or volatiles, such as ethylene, that may inhibit some organisms and stimulate others (Graham and Linderman 1980, Marx 1973). Free-living, nitrogen-fixing bacteria may provide some of the nitrogen required by wood-rotting fungi (Aho and others 1974).
As fallen trees progress from decay class I to class II, the scavengers are replaced by competitors with the enzyme systems needed to decompose the more complex compounds in wood. The fungi involved in this activity are often mutually antagonistic, so that a given part of the tree may be occupied by only one fungus that excludes others by physical or chemical means (Li 1981, Nelson 1967).
The penetration of plant roots through openings in the bark of an advanced class II tree introduces new possibilities for fungal or microbial colonization because the roots offer carbon and nitrogen compounds quite different from those of the wood. Some of these compounds may be secreted from the roots to selectively promote growth of certain micro-organisms (Slankis and others 1964, Smith 1969). Some fungi are capable of penetrating feeder roots to tap the nutrients within. Fungi that damage or kill roots in the process are pathogens. Certain fungi, however, grow on and in plant feeder roots as symbionts that not only acquire photosynthetic products from the host root but also provide nutrients, vitamins, and growth regulators to the host. These symbiotic, fungus-root structures (mycorrhizae) are extremely important in colonization of rotten wood by higher plants (fig. 21).
Page 27
Most plants that grow on rotten wood in old-growth Douglas-fir forests require mycorrhizae to survive; without the mycorrhizal fungi, the host plant cannot acquire adequate nutrients. Western hemlock, spruces, and the Ericaceae are all dependent on mycorrhizae. Hemlock seedlings may survive a year or more without mycorrhizae, but growth is minimal until the mycorrhizal fungi colonize the roots (Christy and others 1982). The fungi must first be introduced into the fallen tree as spores or resting cells;they are transported there by insects, small mammals (Maser and others 1981), wind, rain, or a combination thereof. In a class II tree, the spores need to get through the same openings in the bark used by the plants. They are most likely to be moved through the openings by water or as "hitchhikers" on insects or other arthropods.
Some plants on rotten wood form mycorrhizae when the fungi are available but can also grow without them. Many ferns appear to be in this category (Boullard 1979). A few other plant families, such as the sedge and rush families, rarely form mycorrhizae. None of these are as abundant as the mycorrhizal plants on rotten wood in Douglas-fir forests, perhaps because they are less competitive for the nutrients in rotten wood: Many of the nutrients are in organically bound forms that can be cracked only through action of enzymes not produced by green plants. Mycorrhizal fungi, in contrast, are able to decompose some of these compounds and to translocate the released nutrients to the host (Todd 1979).
As a fallen tree progresses through decay class III, the diversity of plants and associated mycorrhizal fungi may increase, especially if roots grow down through the rotted sapwood into the underlying soil. Sloughing of bark, however, suddenly exposes roots in decayed sapwood to freezing in winter and drying in summer, and many of the fine roots die. As the decayed sapwood sloughs off, plants rooted in it may slough off with it. Even if their roots penetrate partially rotted heartwood underneath, they fall over and often die.
The optimal stage of plant development is attained in the class IV tree; plant roots and mycorrhizae grow through cracks and channels of the brown cubical rotted heartwood. Numbers of species of mycorrhizal and decomposer fungi increase, as does the potential variety of interactions among the fungi and between the fungi and higher plants. The variety of mycorrhizal fungi further increases as lichens and mosses become established on the upper surface of the fallen tree (Gobl 1968). Moss communities capture and accumulate many nutrients from rain, and these nutrients can be cycled into higher plants by the mycorrhizal fungi (Binkley and Graham 1981, Nadkarni 1981). At the same time, fungi and roots that bridge rotten wood to underlying soil may transfer soil nutrients to the fallen tree. As fungal cells or fine roots within the wood die, the minerals acquired from the soil add to the nutrient capital of the tree. These tissues can then be decomposed and their nutrients recycled into further growth of wood-inhabiting organisms.
The class IV tree gradually becomes well permeated with roots as it decays into class V. The well-rotted heartwood provides a moist substrate in which mycorrhizal fungi and roots can continue to interact into the summer, well beyond the time that fungi and fine roots have become dormant or dead from drought in the upper layers of mineral soil (Harvey and others 1978, 1979a; Place 1950).
Page 28
Relatively few species of mycorrhizal fungi seem able to grow in rotten wood compared with the number that grow in soil (Gobl 1968, Kropp and Trappe 1982) perhaps because most mycorrhizal fungi do not produce the enzymes needed to decompose the tannins and other phenolic compounds in wood (Giltrap 1982). Some of the mycorrhizal fungi that inhabit both mineral soil and rotten wood develop much more strongly in the wood than in the soil (Trappe 1962, 1965) and some appear to be restricted to rotten wood (Kropp and Trappe 1982). Most mycorrhizal fungi in rotten wood produce mushrooms or truffles as their spore-bearing, reproductive bodies, but they can do this only if they are in symbiotic association with host roots. This particular result of a fungus-plant interaction occurs mostly in class IV to V trees and is an important feature of the animal-plant interactions discussed in the following sections.
Animal-plant.- The introduction of fungal spores into a recently windthrown, class I tree by bark beetles and wood borers initiates a sequence of complex animal-plant interactions that continues until the tree disappears altogether. Once established and growing under the bark, the fungal colonies are grazed by a variety of invertebrates (McMillen 1976, Petersen 1971, Russell 1979). Many decomposer fungi sporulate within fallen trees and depend on insects for dispersal of spores. Thus, the young bark beetles, emerging from under the bark where they began life as eggs, take with them spores of the associated fungi. Similar relationships between insects and fungi probably occur through all decay stages of fallen trees. Insects eat the fungi and disperse the spores. Many of the fungal grazers are food for predators, so the animal-plant interactions are a prelude to animal-animal interactions.
As the fallen tree progresses through decay classes II and III, the zone of decomposition beneath the bark provides shelter to additional invertebrates and substrate for roots and mycorrhizae. The mycorrhizal fungi are thus introduced to the system as an additional food source to grazers, such as nematodes (Riffle 1971) aphids (Zak 1965) and collembolans (Warnock and others 1982). Slippage of the bark, and eventually of decayed sapwood removes that favorable environment and the organisms within it from the top and sides of the tree; that material, however, does not disappear. Most of it accumulates loosely alongside the log to provide a new habitat favorable to many of the same organisms as before, plus larger animals, such as slugs, snails, salamanders, and small mammals.
Various mites, insects, slugs, and snails feed on the higher plants that become established on rotten wood. These plants also provide cover for the animals, as do the lichens, mosses, and liverworts that colonize fallen trees in decay class IV. Wood-boring beetles, termites, and carpenter ants produce channels in the heartwood that provide passageways for roots. The fruiting bodies of the mycorrhizal fungi, produced from energy supplied by the host plant, can also be a major source of food for insects (Fogel and Peck 1975); other arthropods (Buller 1922); and small mammals, such as the California red-backed vole (Fogel and Trappe 1978; Maser and others 1978a 1978b; Ure and Maser 1982).
Page 29

Collembolan (above)
Specific groups of animals exemplify these generalities. Collembolans are tiny, wingless insects, most of which jump by means of a springlike appendage on the abdomenhence, the common name "springtail." Collembolans living in the soil can be divided into two groups: (1) those that live in the surface layer and feed on fungi, algae, lichens, and decaying vegetation; and (2) those that live deeper in the soil and graze on micro-organisms (Edwards and others 1970, Furniss and Carolin 1977, Zachariae 1962). Both groups probably also exist in fallen trees. Those that feed on fungi, algae, lichens, and detritis could inhabit the protected areas in the exterior bark of a class I tree as their food supply begins to accumulate. As the tree’s interior is made available, first by inner bark feeders and then by wood borers, collembolans gain a new dimension in habitats and niches. By the time a decomposing tree reaches decay class III, insect tunnels have penetrated all levels and the entire tree is "open" so collembolans can live deep within it and graze on such micro-organisms as bacteria.
Because collembolans occupy essentially any atmospheric habitat, they are extremely versatile. The greatest number and variety of species are found in the soil or in contact with it, especially where organic materials abound. Most species are susceptible to desiccation; but in damp environments, species that normally would be restricted to the soil or to litter may occur on the surface of the ground or even in standing trees(Christiansen and Bellinger 1980).
Development of collembolans is not necessarily continuous; growth is interrupted in cold seasons. Development of eggs and immature stages may also be interrupted by dry conditions. Dry periods are survived by a form of diapause, a suspension of growth or development. In a few genera, diapause is even associated with a regressive modification of the mouthparts and digestive tract, and sometimes also with striking external modifications (Christiansen and Bellinger 1980).
Collembolans molt throughout life, from 2 times to over 50 times. Growth apparently continues throughout an individual’s life, but at a decreasing rate after sexual maturity. Sexual activity of adults appears to be restricted to alternate instars (Christiansen and Bellinger 1980).
As with collembolans, the mite fauna diversifies within a fallen tree over time. In fact, the mite fauna in rotten wood may be surprisingly complex. As stated by Krantz (1978,p. 57), "The remarkable diversity in morphology . . . is more than equalled by the variety of behavioral characteristics . . .Specializations in habitat often are paralleled by specializations in structure."

Oribatid mite (above)
Mites can be divided initially into two groups-free living and parasitic. The free-living forms exhibit a variety of food habits. Small-plant eaters include mites that feed on bacteria, algae, fungi, and lichens. Such mites occur in the soil and the litter of the forest floor, and they can inhabit the outer bark of a class I fallen tree as it becomes suitable. As dead plant and animal materials accumulate on a fallen tree, mites (which relish putrid material) become part of the community. The strategy of feeding on dead organisms has been even further refined by the mites; some feed on the decaying material of higher plants (large-plant eaters), whereas others are considered to be general plant eaters, consuming fungi, bacteria, and algae, as well as decaying material from higher plants. The general plant eaters often depend on fungi to soften, decay, and prepare the plant tissues. As organic materials increase, an array of fungi becomes established, and mites that prefer fungi for food appear on the scene (Krantz 1978, Luxton 1972).
Page 30
Mites immigrate into a fallen tree as other organisms, such as wood-boring beetles, penetrate it and modify its habitability. As a fallen tree is colonized by bacteria, fungi, and yeasts, mites that dine on these organisms also take up residence. As these organisms live and die, mites that feed on dead tissues join the community and add to its diversity. Our observations suggest that the mite fauna really begins to flourish as a fallen tree approaches class IV. Finally, as the fauna increases and diversifies, the fecal output generates still another level in the partitioning of food resources available to the mite community.
Even in death, mites and other invertebrates interact with plants. Their empty exoskeletons serve as incubators for the spores formed by certain mycorrhizal fungi (Rabitin and Rhodes 1982).

Common earwig (above)
A larger inhabitant of fallen trees is the common earwig. The earwig is mainly nocturnal and spends the day in cracks and crevices and under the bark of large fallen trees, as well as in other places. Although reputed to feed on tender foliage and dead or decaying vegetable matter, it also devours both living and dead insects (Borror and Delong 1964, Imms 1960, Langston and Powell 1975).
Unlike most insects, the common earwig cares for its eggs and young. After depositing a group of eggs in the soil in winter or early spring, the female rests over them much as a hen does her chicks. The newly hatched earwigs remain under and around their parent until they are able to fend for themselves. Common earwigs become adults in summer, and having wings and being capable of flight, they can disperse. There appears to be one generation per year (Imms 1960, Langston and Powell 1975).
Although some common earwigs may be primary tenants of large fallen trees, others may be secondary or part-time tenants. Nevertheless, they are an important part of the food chain, as will be seen in the next section.
The California red-backed vole is the quintessential example of interactions within fallen trees (fig. 23). The vole heavily uses them for cover; we have found that most class II through class V old-growth Douglas-fir trees have vole tunnels underneath. The vole eats mostly fungi and includes lichens but prefers truffles (Ure and Maser 1982). Some truffles, such as Rhizopogon vinicolor fruit mostly in rotten wood, where their mycelium forms the tuberculate mycorrhizae of conifers (Trappe 1965, Zak 1971) (figs. 24 and 25). The truffle spores are dispersed by animals that eat the truffle (Trappe and Maser 1976). Thus, there is a tight cycle of interdependence: the vole needs the truffle for food; the truffle depends on the vole for dispersal of spores and on a mycorrhizal tree host for energy; the tree requires mycorrhizal fungi for uptake of nutrients and provides the rotten wood needed by the vole for cover. Moreover, since both voles and Rhizopogon vivicolor specialize in rotten wood as habitat, the vole disperses the Rhizopogon spores to the kind of substrate in which the fungus thrives.
Page 31