Abstract
Maser, Chris; Trappe, James M., tech. eds. The
seen and unseen world of the fallen tree. Gen. Tech. Rep. PNW-164. Portland, OR:
U.S. Department of Agriculture, Forest Service, Pacific Northwest Forest and
Range Experiment Station; 1984. 56 p. In cooperation with: U.S. Department
of the Interior, Bureau of Land Management
Large, fallen trees in various stages of decay contribute
much-needed diversity to terrestrial and aquatic habitats in western forests.
When most biological activity in soil is limited by low moisture availability in
summer, the fallen tree-soil interface offers a relatively cool, moist habitat
for animals and a substrate for microbial and root activity. Intensified
utilization and management can deprive future forests of large, fallen trees.
The impact of this loss on habitat diversity and on long-term forest
productivity must be determined because managers need sound information on which
to base resource management decisions.
Keywords: Fallen trees, decay (wood), decomposition,
old-growth stands, Douglas-fir, Pseudotsuga menziesii, mycorrhizae, soil
moisture.
Contents
1 Prolog
1 Introduction
2 The Seen World of the Fallen Tree
2 Recruitment of Fallen Trees
3 Mass of Fallen Trees
4 Placement of Fallen Trees
5 Decay Classes of Fallen Douglas-Fir Trees
7 A Fallen Tree’s Relationship With Time
7 Determining the Age of a Fallen Tree
8 Size and Shape of Fallen Douglas-Fi
10 Substrates as Niches
12 Internal Surface Area
13 Temperature
13 Moisture
15 Plant Rooting
15 Insect Galleries and Frass (Excrement)
16 The Unseen World of the Fallen Tree
17 Microhabitats and Inhabitants
19 Nutrient Cycling
19 Interactions-Nutrients, Plants, and Animals
36 Biotic Succession Within and Around Fallen Trees
38 Stream Characteristics and Fallen Trees
39 A Historical Sketch of Woody Debris in Northwest Waterways
41Vegetation of Streamsides and Gravel Bars
42 Decomposition and Nutrients in Streams
44 Aquatic Invertebrates
46 Fish Habitat
47 Management Impacts on Fallen Trees
47 On Land
48 In Streams
49 Research Needs
49 Epilog
50 Common and Scientific Names
50 Acknowledgments
51 Literature Cited
Prolog
. . dying and dead wood provides one of the two or three greatest resources for
animal species in a natural forest . . . if fallen timber and slightly decayed
trees are removed the whole system is gravely impoverished of perhaps ,more than
a fifth of its fauna.
(Elton 1966, p. 279)
Introduction
Large, fallen trees are unique, critical, dynamic components of old-growth
forests (Franklin and Hemstrom 1980, Franklin and others 1981, Maser and others
1979). Each is a microcosm. Harvey and others (1979b) stated:
Evidence that soil organic reserves, particularly wood, play
important roles in
maintaining forest site quality emphasizes the need to properly manage woody
materials. Thus, the viewpoint that woody residue represents only waste or
a
fire hazard must be reassessed. Forest users and managers must recognize
the benefits, equivalent to long-term fertilization, that woody and other
organic
reserves contribute to forest ecosystems.
Even a casual observer of present western forests would note the abundance of
fallen trees in various stages of decay, whether in a virgin old-growth stand or
a recent clearcut (fig. 1). Up to a century ago western stream systems
also characteristically contained abundant pieces and aggregations of large,
woody debris, but that debris has been systematically removed to improve
navigation, flood control, and drainage. We now have the technological
capability to remove more and more woody debris from the forest floor.
Conversion of forests from virgin to managed status reduces rotation ages from
centuries to decades with a consequent reduction in average size of trees and
change in wood quality. Forests of the future will have far less woody
material contributed to the forest floor than forests of the past, and that
material will differ in size and quality from the woody debris that has been
historically prominent in forest habitats.
Woody debris is generally removed from streams or forests in the name of
economic progress, but what are the short-term and long-term biological
consequences? How is habitat diversity affected, and what is the impact on
long-term site productivity? Our purpose is to encourage awareness and to
facilitate thought on these issues by synthesizing available data on fallen
trees in unmanaged old-growth forests. In so doing, we can also identify
some research needs. The geographic scope is primarily the Douglas-fir
region, but the principles and concepts should apply elsewhere.

Figure 1.-A Douglas-fir recently recruited to the forest
floor.
Page 1
The Seen World of the Fallen Tree
Life on earth, as humans view it, carries but a single
certainty, what lives shall die. Life and death are interdependent. In
forests, this is readily apparent in the large, fallen trees that are a major
component of the forest floor. Although dead, they are an integral part of
the living old-growth forest.
Recruitment of Fallen Trees
Fallen trees are recruited to the forest floor by natural
catastrophic events, such as windstorms that uproot and blow over whole trees or
break their tops (Childs and Clark 1953, Cline and others 1980, Dahms 1949,
Falinski 1978, Roth 1970, Ruth and Yoder 1953) (fig. 2). Heavy snow also breaks
out treetops; and avalanches, mass soil movements, and floods knock down whole
trees (Rothacher and Glazebrook 1968).
Snags-dead, standing trees-on the other hand, usually deteriorate and simply
collapse (Boyce 1923, Boyce and Wagg 1953, Cline and others 1980, Graham 1982,
Keen 1929) (table 1, figs. 3 and 4).

Figure 2.-Fallen trees are natural components of the
forest floor in unmanaged forest.

Figure 3.-Snags are also natural components of the
unmanaged forest.

Page 2
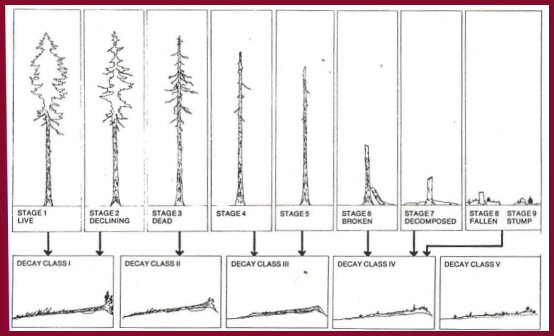
Figure 4.-When they fall, trees and snags immediately
enter one of the first four decay classes (modified from Maser and others 1979).
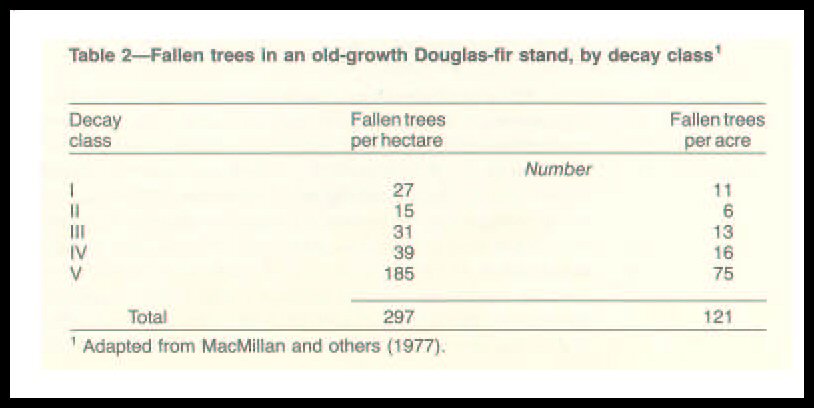
Mass of Fallen Trees
Trees recruited to the forest floor accumulate through time
(table 2, fig. 5); accumulation and rate of decomposition vary with the type of
forest, slope, and aspect (Franklin and others 1981, Grier 1978, Graham and
Cromack 1982, Maser and others 1979). In one midelevation stand of
unmanaged 470-year-old Douglas-fir in western Oregon, the recruitment rate was
estimated to be 0.49 fallen tree per acre (1.2 trees/ha) per year (Grier and
Logan 1978). The decaying Douglas-fir trees represented from 53.4 to 265.4
short tons per acre (120 to 595 metric tons(t)/ha) (MacMillan and others 1977).
Franklin and Waring (1980) showed values ranging from 31.2 to 69.4 short tons
per
acre (70 to 156 t/ha) in stands of widely different ages; their work included
both young stands with large, carryover pieces of rotting wood and old-growth
forests. Grier and Logan (1978) found that as much as 60 percent of the annual
litter fall in a 450-year-old Douglas-fir stand may be woody debris.
Sollins (1982) found that coarse woody debris contributed about 50 percent of
the litter on a long-term basis. Further, decomposing trees in western
Douglas-fir forests represent more volume above ground than is represented by
the aboveground woody debris of typical deciduous forests in the Eastern United
States (Day and Monk 1974, McFee and Stone 1966, Sollins and others 1980)
(fig. 6).
Page 3

Figure 5.-Douglas-fir tree, decay class I, fell before the
Douglas-fir snag, which which entered decay class 3.

Figure 6.-Substantial volumes of decomposing fallen trees
are typical of old growth forests in the Douglas-fir region.
Figure 7.-A fallen tree oriented along the contour of a
slope. The upslope side is filled withy humus and inorganic material that
allows invertebrates and small invertebrates to tunnel alongside. The
downslope side provides protection cover for larger vertebrates.
Placement of Fallen Trees
Fallen trees that are oriented along the contours of a slope
seem to be used more by vertebrates than are trees oriented across contours,
especially on steep slopes. Large, stable trees lying along contours help reduce
erosion by forming a barrier to creeping and raveling soils (fig. 7). Soil
and nutrients deposited along the upslope side of fallen trees reduce loss of
nutrients from the site. Such spots are excellent for the establishment
and growth of vegetation, including tree seedlings.
Vegetation becomes established on and helps stabilize this
“new soil” (Maser and others 1979), and as invertebrates and small vertebrates
begin to burrow into the new soil, they not only nutritionally enrich it with
their feces and urine but also constantly mix it by their burrowing activities.
The interactions of fallen trees with soil are directly
affected by steepness of slope and ruggedness of terrain; a fallen tree on flat
ground, for example, is much more likely to contact the soil over its entire
length than is one oriented either across or along contours on steep or rough
terrain. The proportion of a tree in contact with the soil affects the
water-holding capacity of the wood (Graham 1925). In our studies of fallen
trees in old-growth Douglas-fir forests, the moisture retention through the
summer drought was best in the side of trees in contact with the soil.
The moisture-holding capacity of the wood affects in turn its internal processes
and therefore the succession of plants and animals. In addition, the
orientation of a fallen tree to aspect and compass direction and the amount and
duration of sunlight it receives, drastically affect its internal processes and
biotic community (Graham 1924, 1925; Graham and Knight 1965).
Page 4
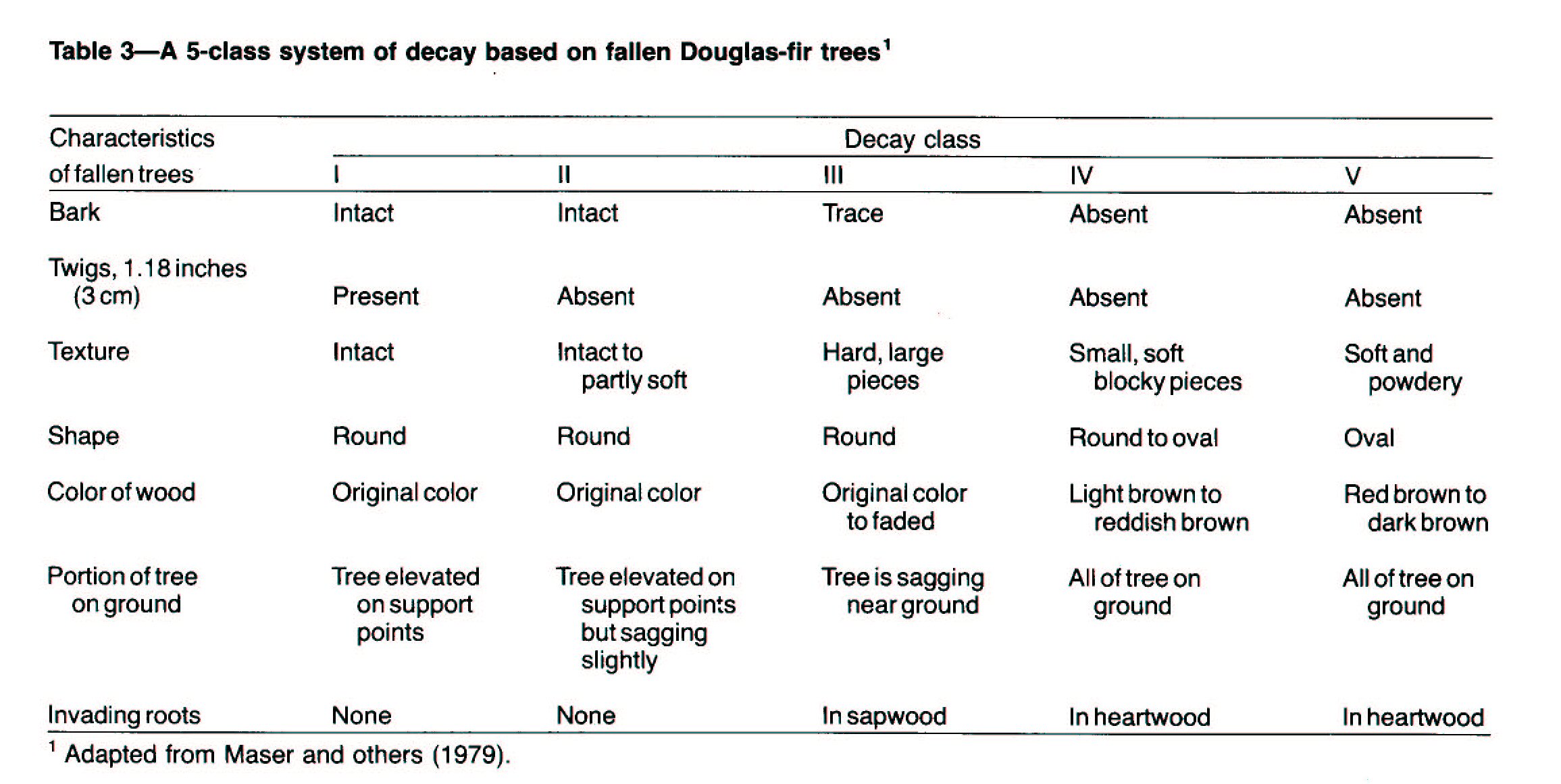
Decay Classes of Fallen Douglas-fir Trees
Dead, fallen trees decay continuously, passing through
recognizable stages or classes of decomposition. Fogel and others (1973)
described broad classes of decay, based on the physical condition of the bark,
wood, and twigs; the presence and pattern of vegetation on a fallen tree and its
degree of root development; and the genera of fungi (identified from fruiting
bodies or sporocarps) associated with the fallen tree. The five decay
classes (Maser and others 1979; table 3), refined by subsequent study (Sollins
1982, Triska and Cromack 1980) are indispensable to research on wood
decomposition for three reasons:
1. These stages of decay are inevitable; despite variation in
the original material and surrounding environment, a classification of decay
based on general processes can be used. (A publication by Minore (1966) can be
used to key fallen trees to species.)
2. Decay class can be used without having to determine when
the tree fell, a difficult problem that requires destructive techniques.
3. The original decay classes form ecological units that
function as distinctive habitats for plants and animals.
The major limiting factor of this decay classification is that
it is based on the external characteristics of a fallen tree and does not
adequately convey the internal diversity of niches. We have found,
however, that internal development of niches relates reasonably well to decay
class.
A 450-year-old Douglas-fir stand on the H. J. Andrews Experimental Forest in the
Willamette National Forest in western Oregon had a distribution of fallen trees
in decay classes II to V that covered an average of 24 percent of the ground
surface, ranging from 11 to 35 percent on different subplots. Although
class V trees produced the most coverage, many were evident only as mossy or
humus-covered mounds on the forest floor. Class I fallen trees, on the
other hand, accounted for the least coverage for two reasons: (1) mortality
rates are low in old-growth stands (Cline and others 1980) and (2) a rapid rate
of decomposition in class I trees quickly converts them to class II.
Subsequent change to higher decay classes progresses more and more slowly as the
most readily decomposed fractions of the wood are utilized by decomposers (Hulme
and Shields 1970).
Variation within and among fallen trees.--One must be aware of two
sources of
variation when using the decay classification: (1) within a particular fallen
tree and (2) among fallen trees of the same decay class.
Variation within a fallen tree.-An old-growth Douglas-fir tapers along
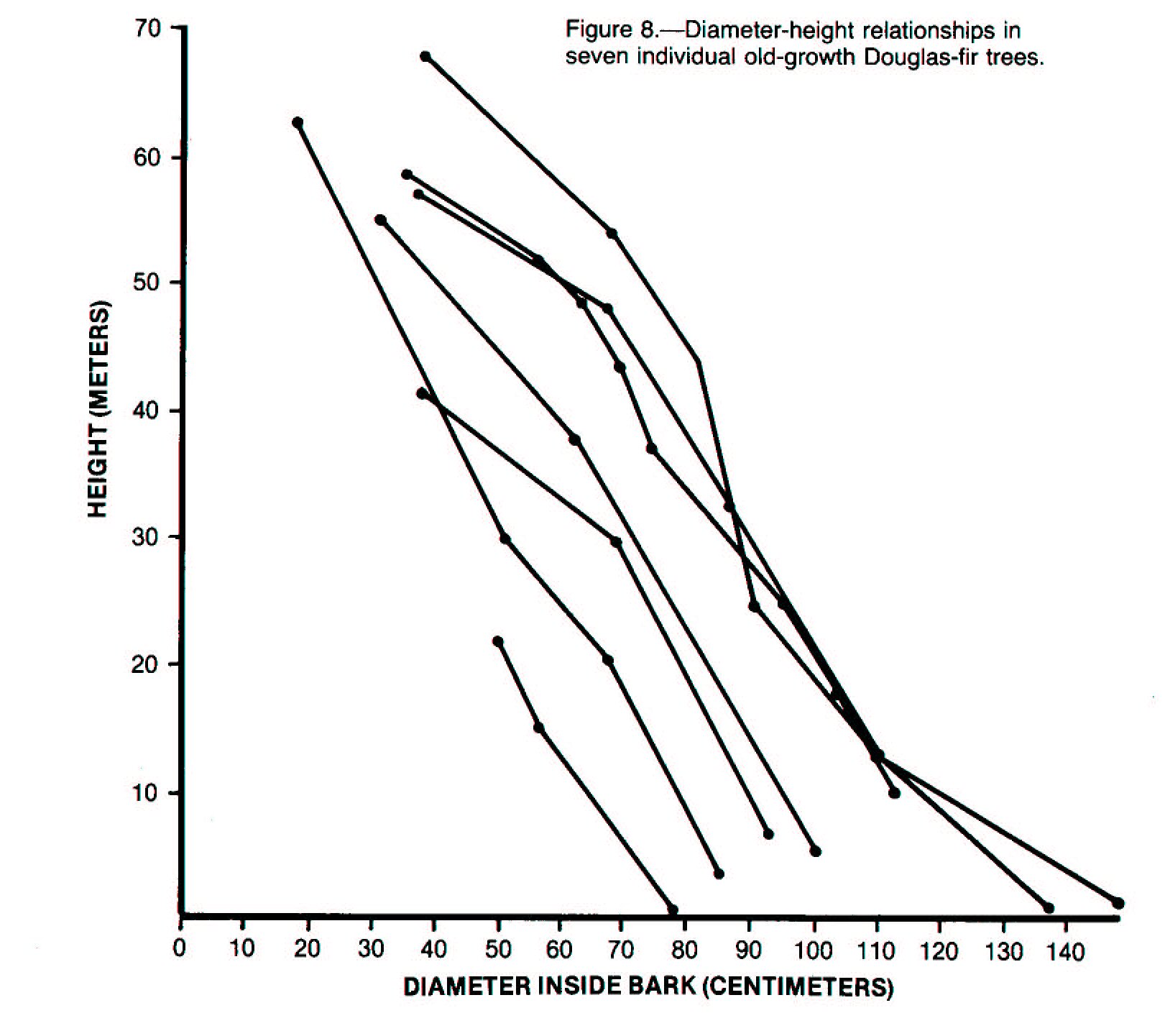
the length of its trunk, from its root collar to the tip of its crown (fig. 8).
Much of the variation within a fallen tree relates to this taper. The
smaller diameter parts of a fallen tree decay faster than the larger ones
because the volume of the wood decreases toward the crown, the ratio of sapwood
to heartwood increases (Dadswell and Hillis 1962), and fragmentation and contact
with the forest floor increase. As decay proceeds, variation within the
tree gradually changes; by the time the tree enters decay class V, the entire
tree becomes a relatively amorphous, homogeneous mass.
Page 5
Page 6
Variation among fallen trees.-The second source of
variation-among fallen trees of the same decay class-is partly a consequence of
classification itself. A single decay class necessarily contains a variety
of fallen trees because they are assigned to a discrete class from a decay
continuum. Additional variation, however, is caused by differences among
trees at the time they fall. For example, trees vary in size, original
diameter, age, old growth versus young growth, decay condition, live tree versus
standing dead tree, and presence of heart rot.
A Fallen Tree's Relationship With Time
The age of a fallen tree-the number of years it lies on the
forest floor-is positively
correlated with decay class, but the relationship is logarithmic rather than
linear. The
time lapse between classes IV and V is exponentially greater than between decay
classes III and IV, and so on. Decay classes I and II evolve rapidly,
whereas the later
classes develop slowly; variation in residence time within a decay class
increases in
the later classes. Estimating the residence time or age of a fallen tree
becomes
increasingly difficult as decay proceeds. Further, decay classes can
evolve at different
rates, depending on the physical setting of a forest stand, such as a
north-facing slope
versus a south-facing slope, or low elevation versus high elevation.
Determining the Age of of a Fallen Tree
The time a fallen tree has rested on the forest floor can be
determined by two standard
methods: (1) aging the scars left on adjacent, living trees that were hit by the
tree when
it fell, and (2) aging the oldest tree growing on the fallen tree (fig. 9).
Scars.-Although aging scars left on the living, woody vegetation by the
tree as it fell
is the most reliable way to age what is now the fallen tree, such scars are
often difficult
to locate because they heal over or are inaccessible. Moreover, the healing
pattern of
wounds varies greatly, so increment cores taken around scars often produce
inaccurate
estimates of residence time.
A more reliable, but destructive, method of aging is to saw either a wedge or a
complete
cross section from the scar area; however, sawing injures or kills trees, an
intolerable
effect on long-term research sites.

Figure 9.-Western hemlock tree and seedlings (background)
growing on a fallen tree that has decomposed into a mound on the forest floor.
Page 7
Seedlings.-The second method of aging a fallen tree is to
age the seedlings growing on it. Aging seedlings is a less reliable method
than is aging scars because the lapse of time between the fall of a tree and the
establishment of the oldest seedling is not known. But this lag can be
calculated for a fallen tree by determining ages for both the scar and the
oldest seedling and then subtracting the latter from the former. Several
pairs of these age counts will reveal the mean lag for the stand. Mean lag
can vary markedly from one stand to another.
Age for fallen trees that did not produce scars but do support
rooted seedlings can be calculated by adding the mean lag to the age of the
oldest seedling. This method of aging fallen trees, however, also requires
the sacrifice of living trees that may be important in the development of a
stand or in future studies.
Size and Shape of Fallen Douglas-Fir
There are three major sources of coarse woody debris in an
old-growth Douglas-fir forest: (1) uprooting of live trees, with or without
complete crowns; (2) breakage of crown and stem of live trees; and (3) breakup
and fall of snags (Graham 1982). Because size of trees and the manner in which
woody material comes to be on the ground vary widely, the resulting pieces of
woody debris are heterogeneous in size and shape. Regardless of its
original size, wood passes through the various stages of decay; the smaller it
is, the faster it breaks down and disappears.
Surveys of large, coarse woody debris in old-growth forests show
that broken, fallen trees are typically more abundant than whole fallen trees
are (fig. 2).¹ To illustrate the
changes in the size and shape of fallen Douglas-fir trees during the
decomposition process, however, we will use a generalized whole, uprooted tree
with a complete crown as an example.
Volume.-The first noticeable decrease in the volume
of a fallen tree is in decay class III (Graham 1982). Bark alone can
account for about 20 percent of the volume of a fallen Douglas-fir (Snell and
Max 1981). As the sapwood is consumed and fragmented by both plants and
animals and sloughs off, about 50 percent of the volume is lost, most of it
during the transition from decay class III to decay class IV. This
transition is called the basic fragmentation stage because of the cubical chunks
of throughly brown-rotted heartwood. Fragmentation continues in decay
class V, but the wood is held together by the prolific rooting of plants (fig.
10). Consequently, only 20 percent of the volume of a decay class V tree
is lost; but with the loss of that 20 percent, only a tenth of the fallen tree’s
original volume remains (Graham 1982).
The bark and wood that slough off a rotting fallen tree
accumulate alongside the tree. This material forms a mulch that extends
over the ground area influenced by the tree.
Diameter.-The diameter of a fallen Douglas-fir decreases
as the outer bark, sapwood, and heartwood slough off. So most of the
overall volume loss is in diameter rather than in length; in our studies, the
transition from decay class III to IV, when the maximum volume is lost,
corresponded to the largest reduction (44 percent) in diameter. By decay
class V, 68 percent of the original diameter has been removed through
decomposition (see footnote 1). Some of the "lost" material is still
present as crumbly fragments of wood and bark on the soil adjacent to the fallen
tree, and some is incorporated into the soil by animal activity. Some has
been physically removed by weather or animals, and some has been used as food by
decomposer organisms.
- - - - - - - -
¹ P. Sollins and S. P. Cline.
Unpublished data on file at Oregon State University, School of Forestry,
Department of Forest Science, Corvallis, Oregon 97331.
Page 8

Figure 10.-Prolific rooting of western hemlock saplings on a
decay class V fallen tree holds the rotten wood together; ground cover is Oregon
oxalis.

A diameter of about 12 inches (30 cm) seems to be a critical
boundary between slow or rapid decay and disintegration of fallen trees.
Pieces smaller than that have a higher proportion of sapwood to heartwood and
disappear more rapidly than pieces of larger diameter. This relationship
between diameter of fallen trees and speed of decomposition has important
ramifications in managed forests in which total tree size will be controlled.
Length.-The length of a fallen Douglas-fir tree decreases
over time from the top toward the roots because the top has the smallest
diameter and decomposes fastest (fig. 11). Little change in length takes
place in decay classes I through III. By the time a tree reaches decay
class IV, its length has been reduced about 15 percent because the top has begun
to be incorporated into the soil. The largest reduction in length (about40
percent) occurs during the transition from decay class IV to V, because the
diameter decreases through fragmentation. Therefore, only a fraction of the
original length of a fallen tree remains by the time it reaches class V (see
footnote 1).
Page 9
Shape-Viewed from above, an intact, fallen tree changes in shape
from a frustum (a truncated cone, tapered at the top) in decay classes I through
III (fig. 5) to nearly cylindrical (both ends with a similar diameter) in decay
classes IV and V. This change in shape can be demonstrated by comparing
the ratio of basal diameter to top diameter: decay classes I and II = 5:l; decay
class III = 4.4:1, decay class IV = 2.1:1; and decay class V = 1.6:1 (see
footnote 1).
In across-sectional view, a fallen tree is circular in decay
classes I through III. By settling and sloughing, it becomes elliptical in
decay class IV. The elliptical shape becomes exaggerated, approaching a
lens shape, in decay class V and later stages.
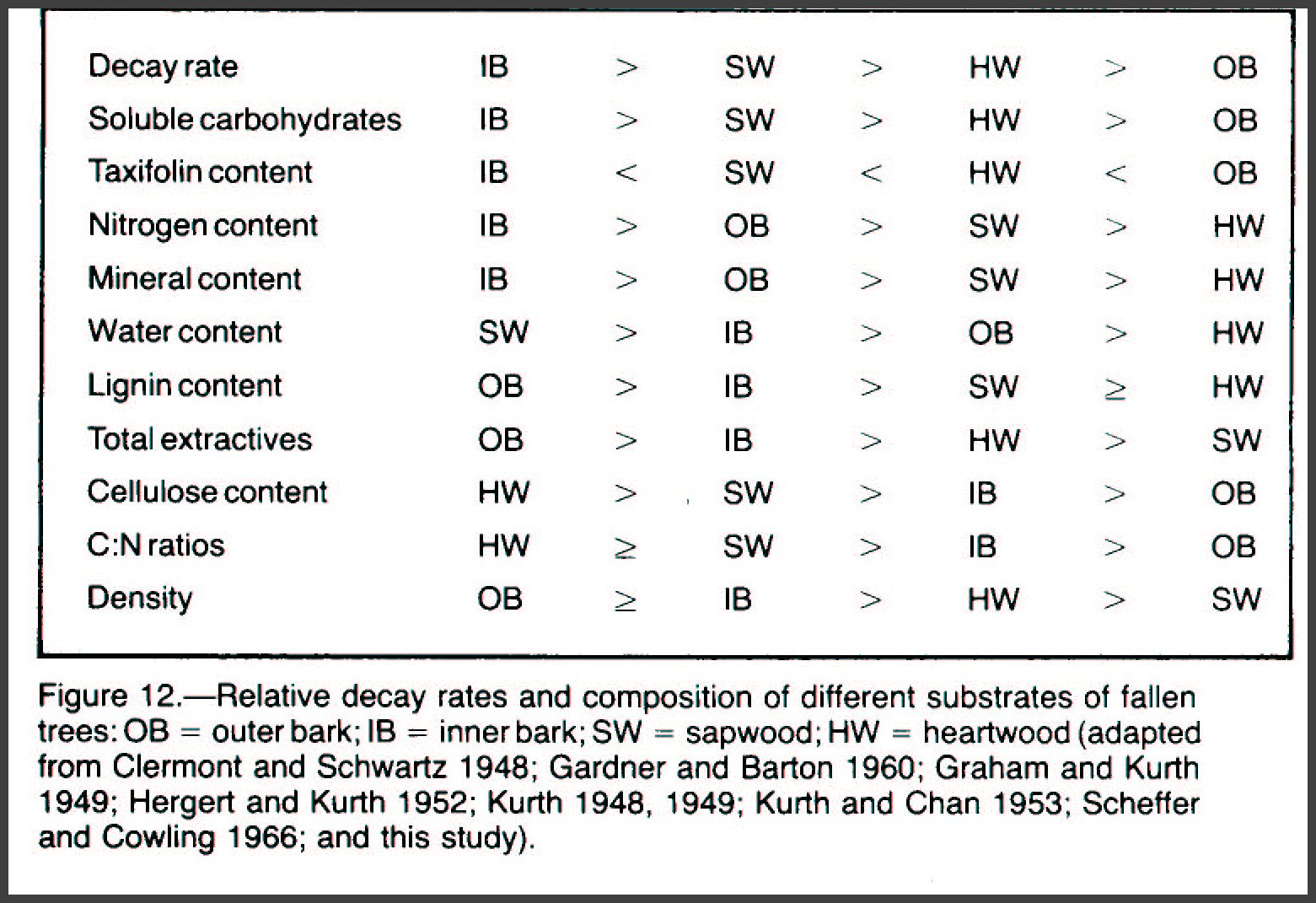
Substrates as Niches Every living conifer is
composed of tissues that perform specific functions. When a tree dies, the
various tissues provide distinguishable substrates that provide different
niches.
The four major tissues of a Douglas-fir tree are outer bark;
inner bark or phloem; sapwood, the living portion of the xylem; and heartwood,
the dead portion of the xylem (fig. 12). A class I fallen Douglas-fir is
mostly xylem, heartwood (60 to 80 percent) and sapwood (5 to 20 percent).
The thick, outer bark of an old-growth Douglas-fir may be 5 to 20 percent of the
cross-sectional area, whereas the inner bark is usually less than 5 percent.
The outer bark and heartwood of a Douglas-fir tree are
physiologically inactive. Outer bark forms a physical and chemical barrier
between the inner tree and the atmosphere, insects, and diseases.
Heartwood supports the tree and stores metabolic wastes (Brown and others 1949,
Hillis 1962). The inner bark and sapwood are physiologically active.
Inner bark is the growing portion of a tree and is the site of both formation of
new cells and transportation of food, whereas sapwood transports and stores
water and dissolved mineral salts (Brown and others 1949).
Page 10
These parts of the fallen tree provide substrates of differing
quality for use by other organisms. Quality of the substrate depends on
physical and chemical properties; the higher the quality, the faster it
disappears through respiration and fragmentation (Lambert and others 1980).
Respiration is the enzymatic transformation by decomposer organisms of organic
compounds to carbon dioxide, water, and other simple products. Fragmentation
occurs when the substrate is eaten, sloughed, and leached.
The main chemical differences among substrates are: (1) nitrogen
content; (2) the mineral or ash content-phosphorus, potassium, calcium,
magnesium; (3) the carbon matrix-cellulose, lignin, pentosans (Lewis 1950); and
(4) the content of other organic compounds-waxes, pigments, carbohydrates, fats,
resins, phenolic compounds (Graham and Kurth 1949, Hergert and Kurth 1952, Kurth
1948).
Many chemical substances are associated with the carbon matrix
but are not chemically bonded to it. Bark contains more such extractable
materials than wood does (Kurth 1948, 1949) (fig. 12); for example, taxifolin, a
natural component of Douglas-fir, has fungicidal properties (Kennedy 1956).
Tissues that were physiologically active in the live Douglas-fir
decay most rapidly after the tree falls because they are higher in quality than
inactive tissues are (fig. 12). Inner bark and sapwood of freshly fallen
trees attract the initial decomposers-beetles that feed on these tissues.
By penetrating the protective outer bark, the beetles open the inner bark and
sapwood to invasion by other decomposers. These tissues contain more
soluble carbohydrates, more moisture, and less taxifolin than do the lower
quality outer bark and heartwood (fig. 12). The substrate of poorest
quality is the decay-resisting outer bark, which is low in moisture,
carbohydrates, cellulose, and carbon to nitrogen (C:N) ratio but high in lignin,
taxifolin, total extractives, and density (fig. 12).
Regardless of substrate, changes develop during decay of fallen
trees: (1) density decreases; (2) water content increases until decay classes
III and IV are reached, at which time the water content stabilizes; (3) mineral
and nitrogen contents increase; 4) the cellulose content decreases; (5) the
relative lignin content increases; and (6) the C:N ratio decreases (Fogel and
Cromack 1977, Foster and Lang 1982, Graham and Cromack 1982, Grier 1978, Lambert
and others 1980, Sollins 1982).
Inner bark disappears fastest because it has the highest
substrate quality and the smallest volume. Both sapwood and outer bark
disappear by decay class IV, but for different reasons. Sapwood elicits
high biological activity and disappears because of insect consumption and
microbial breakdown (fig. 13). Outer bark disappears almost solely by
fragmentation and sloughing from the top and sides of a fallen tree. Large
trees are mostly heartwood when they fall and by decay class IV only heartwood
remains; without the initial association with higher quality substrates, such as
sapwood and inner bark, heartwood would undoubtedly decay more slowly (see
footnote 1). When decay reaches the advanced class V stage, the fallen
tree appears as a mound on the forest floor, usually covered with humus and
litter or moss and often supporting several to many hemlocks. Much of the
crumbly, brown-rotted heartwood remains in place. That which has
disappeared, however, is not all lost through metabolism of decomposers.
Much of it merges into the humus or becomes incorporated into the soil profile
(Denny and Goodlett 1956; Harvey and others 1978, 1979b; McFee and Stone 1966).
Page 11

Figure 13.-The inner bark and part of the
sapwood of this decay class II fallen tree have decomposed; the outer bark and
heartwood are still intact.

Figure 14.-A fallen Douglas-fir tree in decay
class IV, with wood separated into the characteristic angular blocks known as
brown cubical rot; this decay stage provides superb rooting substrate for
hemlock seedlings.
Internal Surface Area
The importance of internal surface areas is that, through such
surfaces, a fallen tree interacts with its environment. A newly fallen
tree, for example, interacts only passively with the surrounding forest because
its interior is not accessible to plants and most animals. But once fungi
and bacteria, which are smaller than the wood fibers, gain entrance, they slowly
dissolve and enter the wood cells. And wood-boring beetles and termites
chew their way through the wood fibers. Meanwhile, many other organisms,
such as plant roots, mites, collembolans, amphibians, and small mammals, must
await the creation of the internal spaces before they can enter. The flow
of plant and animal populations, air, water, and nutrients between a fallen tree
and its surroundings increases as the aging process continues.
Surface area within a fallen tree develops through physical and
biological processes. A tree cracks and splits when it falls and
subsequently dries. Microbial decomposition breaks down the material in
the cell wall and further weakens the wood. Wood-boring beetle larvae and
termites tunnel through the bark and wood; this activity not only inoculates the
substrates with microbes but also opens the tree to colonization by still other
microbes and small invertebrates. Wood-rotting fungi produce zones of
weakness, especially between annual rings, by causing spring wood to decay
faster than summer wood; and plant roots that penetrate the decayed wood split
it as the roots elongate and thicken in diameter. Because of all this
internal activity, the longer a fallen tree rests on the forest floor, the
greater the development of its internal surface area. Most internal
surface area results from biological activity, the cumulative effects of which
not only increase through time but also act synergistically-insect activity
promotes decomposition through microbial activity that encourages the
establishment and rooting of plants.
Most splitting of the sound wood found in decay classes I-III in
fallen trees is radial as the wood dries. Later, the weakening of annual
rings by the more rapid decomposition of spring wood than summer wood leads to
circumferential cracking as well. The blocky structure in class IV and V
fallen trees results from brown cubical rot (fig. 14).
Page 12
Temperature
Gross environmental features regulate temperature regimes.
Latitude, elevation, aspect, and vegetation not only set the maximum and minimum
temperature of a site but also control the seasonal range in temperature.
For fallen trees, the question is twofold: (1) What is the relationship between
the temperatures of air, tree, and soil? and (2) How is this relationship
modified through the year by characteristics of the tree itself?
Summer data (see footnote 1, p. 8 ) indicate the following
temperature relationships at all elevations sampled: air : fallen tree > soil.
The absolute differences were greater at low elevations than at midelevations or
high elevations; mean winter temperatures of the air, a fallen tree, and the
soil were nearly identical at each elevation.
Substrate temperatures within fallen trees of a particular decay
class differ little at a given time and site, but each decay class responds
differently to the temperatures of the surrounding air and soil in summer.
Measurements (see footnote 1, p. 8 ) in August showed the following relationship: air = I = II>III>IV=V>soil. These relationships develop because, as decay proceeds, a fallen tree more closely
hugs the soil, which buffers it against fluctuations in air temperature. Thus, trees
in decay classes I and II are cylindrical and contact the soil relatively little. Trees in
class III have sloughed and slumped somewhat, and those in classes IV and V are partially to
mostly embedded in the soil.
Moisture
Whole fallen tree.-Three trends are visible in the
water-holding capacity of a fallen tree; the same is true for individual
substrates -outer bark (OB), inner bark (IB), sapwood (SW), and heartwood (HW):
1. Water content increases with residence time on the forest
floor and with stage of decay (fig. 15, sapwood) because of the microbial
breakdown of woody substrates that produces water and carbon dioxide; also,
microbial activity increases as decay advances. Simultaneously, the
water-holding capacity of woody substrates increases as cell walls break down.
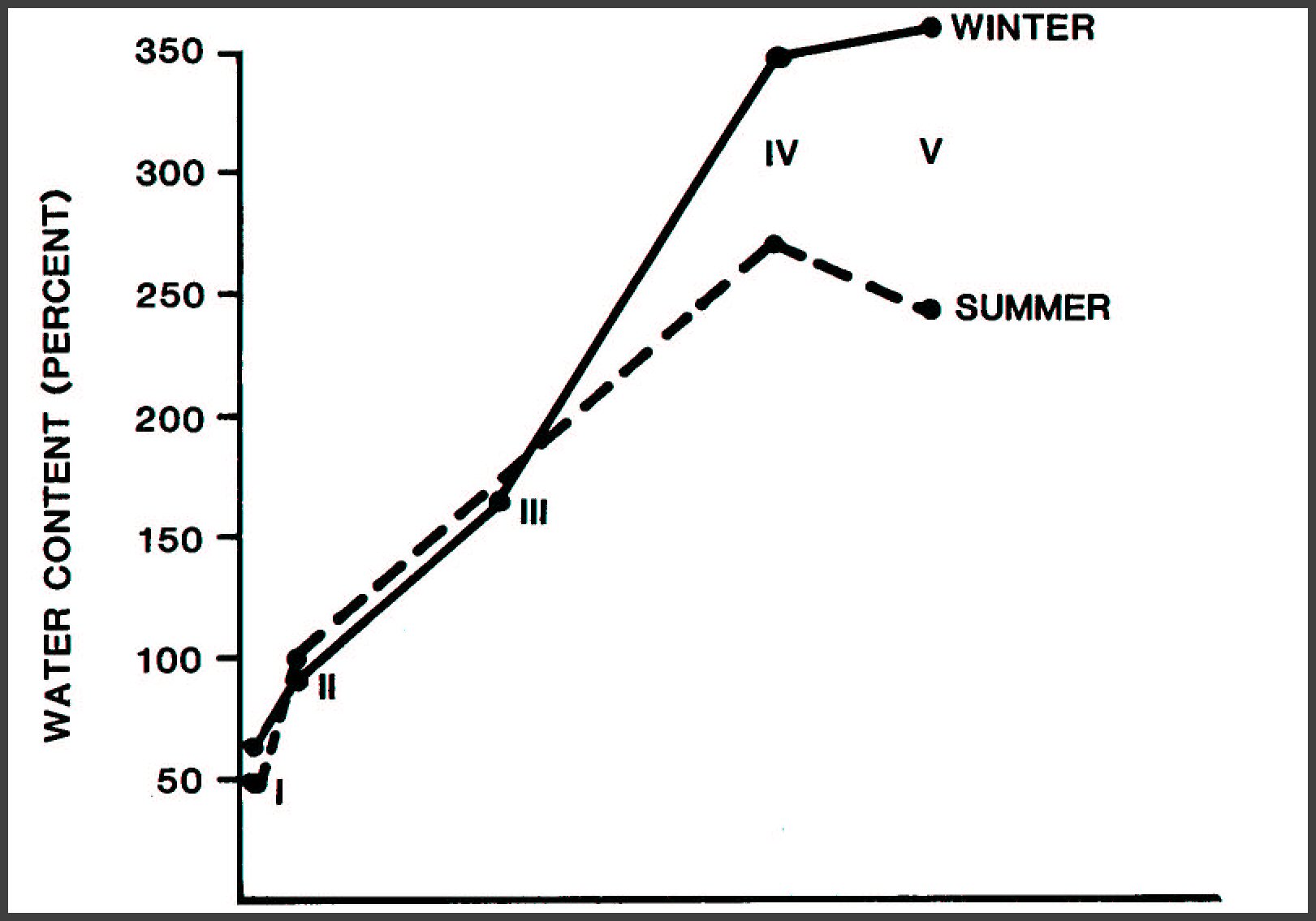
Figure 15.-Water content of fallen Douglas-fir trees in
winter and summer; each point represents a decay class from I (newly fallen
trees) to V (fallen trees decomposed to a mound of brown cubical rotted wood).
Page 13
2. In decay classes I through III, the mean water content of
fallen trees is nearly equal in summer and winter; but in decay classes IV and
V, trees hold more water in winter than in summer. There are two reasons:
(1) Winter usually provides abundant water, so the complete storage capacity of
the woody substrates is used; and (2) evaportranspiration potential is normally
low because days are short, temperatures are cool, and humidity is high.
3. As the overall water-holding capacity of woody substrates
increases with stage of decay, so does the seasonal fluctuation of the water
content (fig. 15).
Substrates. -Four trends are visible in the water-holding
capacity of the substrates within a fallen tree:
1. When a tree falls (decay class I), the water content of the
substrates is: IB>SW>OB>HW. Initially, the inner bark and sapwood (both
physiologically active (alive) at the time the tree fell) contain more water
than do the outer bark and heartwood (both physiologically inactive (dead) at
the time the tree fell). Nearly all of the living cells are located
in the inner bark and sapwood of a live tree.
2. The water content of all woody substrates increases during
the residence time of a tree and its stage of decay. Again, as with the
whole tree, breakdown of woody substrates by microbial activity produces water
and carbon dioxide, and microbial activity increases as decay proceeds.
3. The water content of substrates increases at different rates
within and among seasons. In winter, for example, water content increases
(percent per year): IB>SW>HW>OB. The water retention capacity depends on a
given substrate’s stage of decay; the more advanced the decay, the more porous
the wood and the more water it retains. In summer, however, water-holding
capacity shifts in response to exposure of the substrate to evapotranspiration.
A fallen tree dries because of the dramatic increase in evapotranspiration
brought about by the simultaneous increase in day length and air temperature and
decrease in rainfall and relative humidity. The inner bark, for example,
is protected from excessive drying because inner bark is sandwiched between the
thick outer bark and the relatively moist sapwood And, being the most
nutrient-rich substrate, it supports the highest microbial activity, through
which additional water is produced. Heartwood retains water because of its
large volume; thus, although the exposed outer surface of a tree may dry, the
center retains moisture. Microbial activity continues in the moister areas
and produces more water. Outer bark, on the other hand, is prone to drying
not only because it is exposed to direct sunlight and wind but also
because microbial activity is low, so little water is produced. As the
outer bark sloughs off, the sapwood is no longer protected from sun and wind, so
it begins to dry out. The drying of sapwood is speeded up because sapwood
is more porous than outer bark; therefore, the ability of sapwood to retain
water against the forces of evapotranspiration decreases as porosity increases.
4. Seasonal fluctuations of water content increase as the
water-holding capacity of the woody substrate increases (fig. 15).
Page 14
Plant Rooting hereiam
Woody plants do not root in fallen trees in decay class I.
A tree in decay class II may have plants rooting in its inner bark, which decays
fastest of all the substrates; these roots would have reached the inner bark
through fissures in the outer bark. By the time a tree reaches decay class
III, its inner bark is completely decomposed, and the outer bark and sapwood are
often penetrated by roots (in 50 percent of our samples).
Only about a third of the heartwood samples in decay class III trees were
colonized by roots. When a tree reaches decay class IV, however, only
heartwood remains, and nearly all our samples (87 percent) contained roots.
Finally, trees in decay class V are not only completely colonized by roots but
are actually held together by them (see footnote 1, p. 8).
Within a decay class, plant rooting differs among substrates for
two reasons:
(1) Substrates decay at different rates, and (2) substrates are not equally
accessible to plant roots. In general, as decay proceeds, plant rooting
increases in all substrates because they become excellent rooting medium-density
and hardness decrease, and water content increases.
Insect Galleries and Frass
(Excrement)
Wood-boring insects, such as beetles, carpenter ants, and
termites, tunnel within fallen Douglas-fir trees and consume the woody tissues.
Their activities are evidenced by their galleries and tunnels often packed with
borings and feces. The collective activity of such wood-boring insects
appears to be the most important factor in initiating early decomposition of
fallen trees-decay classes I-III-for three major reasons.
First, wood is broken down by consumption and digestion by
insects and by enzymatic attack by microbes. Insects, however, can
penetrate the wood matrix faster than microbes can. Second, consumption
and digestion of wood also fragment it. As a fallen tree is fragmented by
insect tunnels and galleries, its internal volume is opened to decomposer plants
and animals. The wood matrix is reduced to particles (borings and feces),
and more surface area is available for microbial attack (Crossley 1976).
Third, wood-boring insects serve as vectors for decomposer micro-organisms, such
as intestinal inhabitants that are expelled in the feces (Breznak 1982). They
are also
introduced as external associates; fungal spores, for instance, are transported
in special
pits and cavities in adult beetles (Francke-Grosmann 1967). Fungal spores and
hyphae
are also picked up and carried by insect larvae. In addition, each species of
insect that
tunnels in Douglas-fir bark or wood has its attendant predators, parasites, and
scavengers (Deyrup 1981).
Frequency of galleries by decay class of tree.-Studies in the H. J. Andrews
Experimental Forest in western Oregon showed that the percentage of wood samples
with galleries increased as decomposition of the fallen trees proceeded (see
footnote
1, p. 8). When the substrates within a decay class were compared for galleries,
they
ranked as follows: IB>SW>HW. Outer bark was not included in this comparison
because
the nature of the galleries differed. Insects simply chew through, not within,
the outer
bark to gain access to the nutritious inner bark and sapwood Inner bark
disappeared
before the frequency of galleries reached 100 percent. But all the sapwood
samples
had galleries by the time they reached decay class III and the heartwood, by
decay
class V.
Effects of galleries on wood properties.-Samples of a substrate with insect
galleries
in a fallen tree were softer and wetter and had a lower density, more plant
roots, higher
microbial activity, higher nitrogen fixation activity, higher microarthropod
populations,
and higher exchangeable and mineralizable nitrogen than did samples from the
same
substrate without insect galleries (see footnote 1, p. 8).
Page 15
Back to Main Page